Jedną z cech komórek żywych jest ich pobudliwość, która umożliwia wprowadzenie komórki w stan pobudzenia.
Pobudliwość jest zdolnością komórki do odpowiedzi na bodziec. Tkanki zbudowane z komórek pobudliwych, mające zdolność do szybkiej reakcji na bodźce, nazywamy tkankami pobudliwymi. Zaliczamy do nich tkankę mózgową i nerwy zbudowane z komórek nerwowych, oraz tkankę mięśniową gładką, poprzecznie prążkowaną oraz sercową zbudowaną z komórek mięśniowych. Bodźce np. w postaci krążących w przestrzeni zewnątrzkomórkowej substancji chemicznych mają zdolność wiązania się ze specyficznymi dla siebie receptorami. To połączenie może skutkować:
- otwarciem kanałów jonowych, co umożliwia przepływ odpowiednich jonów przez błonę komórkową;
- aktywacją enzymów znajdujących się w błonie komórkowej.
Odpowiedź na bodziec w postaci w/w zmian właściwości błony komórkowej zachodzi bardzo szybko- w ułamku sekundy. Natomiast następujące po niej zmiany w metabolizmie komórki zachodzą znacznie wolniej.
Pobudzeniem określa się zmianę we właściwościach błony komórkowej lub metabolizmu komórki, która jest wynikiem zadziałania na komórkę czynników zewnętrznych -bodźców. Największą grupę bodźców stanowią różnego rodzaju substancje chemiczne. W organizmie istnieją również zgrupowania komórek pełniące funkcję narządów odbiorczych- receptorów, które mają zdolność reagowania na bodźce fizyczne, takie jak ciepło czy energia mechaniczna. Zarówno bodziec fizyczny jak i chemiczny o odpowiednio dużym natężeniu, może wywołać pobudzenie komórki. Bodziec fizjologiczny to taki bodziec, który wywołuje całkowicie odwracalne pobudzenie komórki, nie powodując przy tym jej uszkodzenia.
Błonowy potencjał spoczynkowy
Cechą komórek pobudliwych jest posiadanie stałego, ujemnego, spoczynkowego potencjału błonowego. Jego wartości podane są w tabeli.
|
Rodzaj komórki
|
Wartość potencjału błonowego
|
|
Neuron
|
-60mV do -80mV
|
|
Komorka mięśnia poprzecznie prążkowanego
|
-80mV do -90mV
|
Płyny wewnątrz- i zewnątrzkomórkowy różnią się istotnie pod względem stężenia w nich poszczególnych jonów. W płynie wewnątrzkomórkowym przeważają organiczne aniony, które nie mają zdolności przechodzenia przez błonę komórkową. Błona komórkowa ulega więc polaryzacji: po jej wewnętrznej stronie występuje ładunek ujemny- wynikający z nagromadzenia anionów, natomiast na zewnątrz komórki- ładunek dodatni.
 rys. Polaryzacja błony komórkowej: "+"- ładunek dodatni; "-" - ładunek ujemny
rys. Polaryzacja błony komórkowej: "+"- ładunek dodatni; "-" - ładunek ujemny
Jeśli metabolizm komórki się nie zmienia i nie działają na nią żadne bodźce, ilość jonów nagromadzona w jej wnętrzu jest stała. Pojawia się wówczas stan pewnej dynamicznej równowagi między stężeniami jonów wewnątrz i na zewnątrz komórki. Przez błonę komórkową ustala się gradient stężenia oraz gradient ładunków elektrycznych poszczególnych jonów.
Stosując równanie Nernsta, które opisuje dyfuzję cząstek naładowanych, wyliczono potencjał wnętrza komórki mięśnia poprzecznie prążkowanego. Wynosi on -90mV.
Jako, że przepływ jonów - powodujący powstanie prądu jonowego- przebiega zarówno do wnętrza komórki jak i w kierunku przeciwnym, przyjmuje się, że w błonie komórkowej istnieją oddzielne kanały dla przepływu każdego z jonów. Otwieranie i zamykanie kanałów jonowych zależy bezpośrednio tego w jakim stanie czynnościowym znajduje się błona komórkowa. Kanały jonowe zbudowane są z białek będących składowymi błony komórkowej, a ich wzajemny układ przestrzenny decyduje o tym, czy kanał jest otwarty czy zamknięty.
Pompa Na+↔K+
Aby utrzymać wysokie stężenie jonów potasowych oraz niskie stężenie jonów sodowych wewnątrz komórki konieczny jest aktywny transport tych jonów przez błonę komórkową wbrew gradientowi ich stężeń.
Enzym zwany ATP-azą sodowo-potasową zapewnia odpowiednie rozmieszczenie jonów i utrzymanie ich gradientu przez błonę komórkową. Cząsteczka enzymu, o łącznej masie ok. 294kD, zbudowana jest z czterech podjednostek: dwóch α (m.cz. 92kD) i dwóch β (m.cz. 55kD). Jony sodowe zgodnie z gradientem wnikają do wnętrza komórki przez kanały sodowe. Jednak wbudowana w błonę komórkową ATP-aza sodowo potasowa, wiąże jony sodu i wypompowuje je na zewnątrz. Jednocześnie po zewnętrznej stronie błony komórkowej enzym przyłącza jony potasu i wpompowuje je do wnętrza komórki.
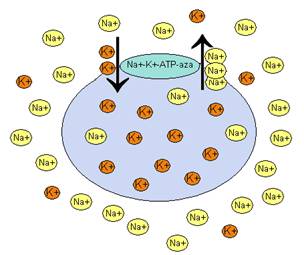 rys. Działanie pompy sodowo-potasowej
rys. Działanie pompy sodowo-potasowej
Energia konieczna do przenoszenia jonów wbrew ich gradientowi pochodzi z rozkładu ATP do ADP. Hydroliza ATP zachodzi pod wpływem jonów sodowych i potasowych i stąd pochodzi nazwa enzymu (Na+-K+-ATP-aza).
Pompa sodowa potasowa działa na zasadzie antysportu tzn. że przenosi jony sodu i potasu transportowane są w przeciwną stronę. Energia pozyskana z hydrolizy jednego mola cząsteczek ATP wystarcza na przeniesienie trzech moli Na+ i dwóch moli K+. Do rozkładu ATP konieczna jest również obecność jonów Mg2+ w płynie komórkowym.
Właściwa praca pompy sodowo- potasowej ma kluczowe znaczenie dla zachowanie polaryzacji błony komórkowej, a tym samym pobudliwości komórki. Na ten cel pozostająca w stanie spoczynku komórka pobudliwa przeznacza około 1/3 swojego metabolizmu. Do prawidłowego działania tych mechanizmów konieczne jest:
- odpowiednie zaopatrzenie komórki tlen i substancje odżywcze (np. glukozę)
- resynteza ATP z ADP zachodząca w procesie oddychania wewnątrzkomórkowego
- usuwanie produktów przemiany materii - głównie dwutlenku węgla
- utrzymanie odpowiedniego stosunku stężeń jonów sodowych do potasowych w płynie pozakomórkowym
- utrzymanie temperatury w komórce na poziomie 37o C, która zapewnia właściwe funkcjonowanie wewnątrzkomórkowych układów enzymatycznych.
Dla prawidłowego funkcjonowania pompy, wszystkie te parametry muszą zostać utrzymane na właściwym poziomie. Zmiana jednego z nich (np. niedotlenienie komórki, spadek temperatury tkankowej) może prowadzić do zatrzymania lub zwolnienia jej pracy. Kiedy pompa przestaje działać dochodzi do wyrównania stężeń jonów po obu stronach błony komórkowej, co w konsekwencji prowadzi do zaniku potencjału spoczynkowego błony komórkowej. W komórkach mięśniowych proces ten zachodzi dość wolno - potrzeba kilkudziesięciu minut, natomiast w komórkach nerwowych wystarczy kilka sekund. Komórki pobudliwe tracą wówczas swoje właściwości i nie są zdolne do reakcji na bodźce. Stają się komórkami niepobudliwymi.
Nieprawidłowa praca pompy sodowo- potasowej prowadzi do zmian nie tylko w składzie płynu wewnątrzkomórkowego ale także prowadzi do zmiany stężenia jonów w płynie zewnątrzkomórkowym. Dzieje się tak dlatego, że tkanki pobudliwe stanowią duży odsetek naszej masy ciała. W konsekwencji w płynie zewnątrzkomórkowym maleje stężenie jonów sodowych, a wzrasta stężenie jonów potasu.
Komórki nerwowe
Ocenia się, że w ludzkim ciele znajduję się 1 bilion komórek nerwowych, wpieranych przez kilkanaście razy liczniejsze komórki glejowe. Najwięcej neuronów skupionych jest w centralnym systemie nerwowym. Ich podstawową funkcją jest przewodzenie impulsów nerwowych, które kodują informacje, płynące do i z naszego mózgu. Przewodzenie impulsów nerwowych jest możliwe dzięki licznym procesom elektrochemicznym, które zachodzą w błonie komórkowej neuronu.
Budowa neuronów jest bardzo zróżnicowana zarówno pod względem ich wyglądu jak i pełnionych funkcji. Jednak w każdym neuronie można wyróżnić ciało komórkowe i jego wypustki : akson i dendryty. W ciele komórkowym zachodzą wszelkie procesy syntezy i metabolizmu komórki. Komórki podporowe otaczają ciało komórki. Ze względu na pełnione funkcje dzieli się ja na komórki makro- i mikroglejowe.
Długość aksonów komórek nerwowych jest bardzo różna. W ludzkim ciele można wyróżnić neurony z krótkimi aksonami, rozgałęziającymi się zaraz po odejściu z ciała komórki oraz neurony z długimi aksonami, o długości nawet 1,2 m. Jądra ruchowe pnia mózgu i rogów przednich rdzenia kręgowego zbudowane są z największych neuronów, z których odchodzą długie aksony.
Potencjał czynnościowy
Wynikiem zadziałania bodźca na błonę komórkową, jest powstanie potencjału czynnościowego. Dzięki otwierającym się kanałom sodowym do wnętrza neuronu dostają się jony sodu, co prowadzi do wyrównania gradientu ładunku elektrycznego przez błonę komórkową. Jest to depolaryzacja błony komórkowej. Początkowo napływ jonów sodu do wnętrza komórki zachodzi tylko w miejscu działania bodźca. Zniesienie gradientu ładunków w tym miejscu rozszerza się następnie na sąsiadujące z nim dalsze odcinki błony komórkowej w postaci tak zwanej fali depolaryzacji. Szerząca się wzdłuż błony aksonów fala depolaryzacyjna to impuls nerwowy. Biegnie on od miejsca, w którym zadziałał bodziec, do zakończenia wypustki neuronu.
Synapsa
Między komórkami nerwowymi impulsy nerwowe są przekazywane w miejscu styku zakończeń aksonów i dendrytów. Synapsa to połączenie zakończeń dwóch wypustek nerwowych, miejsce w którym stykają się ich błony komórkowe i następuje przekazanie impulsu nerwowego z jednej komórki do drugiej. Synapsę budują:
- błona presynaptyczna- zakończenie wypustki nerwowej przekazujące impuls
- błona postsynaptyczna - zakończenie wypustki nerwowej odbierające impuls
Między błonami znajduje się niewielka przestrzeń, która tworzy szczeliną synaptyczną.
Końcowy odcinek aksonu, który tworzy synapsę ulega poszerzeniu tworząc tzw.: synaptyczną kolbę końcową, pokrytą błoną presynaptyczną. Kolby końcowe pokrywają powierzchnię ciał komórkowych i dendrytów, gdzie tworzą liczne połączenia synaptyczne.
Kolby synaptyczne zawierają liczne mitochondria i wypełnione są pęcherzykami synaptycznymi, zawierającymi neurotransmitery i inne substancje chemiczne modulujące przekaźnictwo synaptyczne. Zawartość pęcherzyków synaptycznych zostaje uwolniona do szczeliny synaptycznej, gdy do kolby synaptycznej dociera impuls nerwowy, powodując depolaryzację błony presynaptycznej. Znajdujące się w przestrzeni szczeliny synaptycznej neurotransmitery ulegają związaniu ze specyficznymi dla siebie receptorami.
Im częściej dana synapsa przewodzi impulsy nerwowe, tym większe zagęszczenie pęcherzyków synaptycznych wewnątrz kolby końcowej i tym więcej receptorów dla neurotransmiterów pokrywa błonę pre- i postsynaptyczną synapsy. W synapsach, które rzadko przewodzą impulsy nerwowe zagęszczenie tych struktur jest mniejsze.
Synapsy pobudzające
Z zakończeń presynaptycznych błony komórkowej aksonów do szczeliny synaptycznej uwalniane są różne substancje chemiczne nazywane neurotransmiterami. Docierają one do błony postsynaptycznej powodując zmianę jej właściwości.
Jeśli neurotransmiter doprowadzi do zmniejszenia potencjału błony postsynaptycznej wywołując jej depolaryzację, wówczas taki rodzaj połączenia między komórkami nerwowymi nazywamy synapsą pobudzającą. Połączenie neurotransmitera z błoną postsynaptyczną powoduje napływ jonów sodu do wnętrza komórki i obniżenie się ujemnego potencjału panującego wewnątrz komórki nerwowej, co wywołuje powstanie postsynaptycznego potencjału pobudzającego.
Synapsy hamujące
Jeśli z zakończenia presynaptycznego uwalniany jest neurotransmiter, który powoduje hiperpolaryzację (nadmierną polaryzację) błony komórkowej, wówczas takie połączeni komórek nerwowych nazywamy synapsą hamującą.
Do nadmiernej polaryzacji błony komórkowej dochodzi na skutek zwiększenia ujemnego potencjału panującego we wnętrzu komórki. Jest to spowodowane działaniem neurotransmitera na błonę postsynaptyczną, który powoduje otwarcie kanałów chlorkowych i napływ do wnętrza komórki ujemnie naładowanych jonów chlorkowych. W ten sposób zwiększony zostaje ujemny potencjał panujący we wnętrzu komórki. Jest to postsynaptyczny potencjał hamujący. Błona komórkowa ulega hiperpolaryzacji co prowadzi do obniżenie pobudliwości komórki.
Przekazywanie impulsów nerwowych między neuronami zachodzi stale, a jeden neuron może tworzyć synapsy zarówno pobudzające jak i hamujące.
Neurotransmitery pobudzające
Do neurotransmiterów pobudzających zaliczamy te substancje chemiczne, które docierając do błony postsynaptycznej powodują jej depolaryzację. Ich połączenie z receptorem w błonie postsynaptycznej powoduje powstanie tzw.: aktywacji sodowej czyli otwarcia kanałów dla dokomórkowego przepływu jonów sodowych.
Najważniejsze z nich to: acetylocholina, dopomina, noradrenalina, serotonina oraz aminokwasy pobudzające (asparaginiany i glutaminiany).
Około 75% pobudzającego przekaźnictwa synaptycznego w obrębie centralnego systemu nerwowego zachodzi z udziałem aminokwasów pobudzających. Poza rolą w przekazywaniu impulsów nerwowych wykazują one także negatywny wpływ na neurony. W sytuacji ostrego niedotlenienia tkanki mózgowej ich nagłe uwalnianie z pęcherzyków synaptycznych, powoduje uszkadzający, cytotoksyczny wpływ na neurony.
Innym ważnym przekaźnikiem jest tlenek azotu (NO), pełniący wiele różnych funkcji w całym organizmie. W obrębie synapsy jest on uwalniany przez błonę postsynaptyczną a miejscem jego wychwytu są receptory błony presynaptycznej. To wsteczne działanie NO na błonę presynaptyczną, ułatwia uwalnianie zawartego w kolbie końcowej neurotransmitera do szczeliny synaptycznej.
Neurotransmitery hamujące
Do neurotransmiterów hamujących zaliczamy te substancje chemiczne, które docierając do błony postsynaptycznej powodują jej hiperpolaryzację. Ich połączenie z receptorem w błonie postsynaptycznej powoduje powstanie tzw.: postsynaptycznego potencjału hamującego.
Najważniejszym z nich jest kwas γ-aminomasłowy (GABA). Mechanizm działania GABA na komórkę różni się w zależności od tego z jakim receptorem zostanie om związany. Wśród receptorów GABA wyróżniamy receptory GABAA i GABAB. Połączenie receptorem GABAA powoduje otwarcie kanałów chlorkowych i dokomórkowy napływ ujemnie naładowanych jonów chlorkowych. Receptor GABAB zaliczany jest do receptorów metabotropowych co oznacza, że jego działanie wynika z uruchomienia układu wtórnych przekaźników. Do hiperpolaryzacji błony komórkowej dochodzi wówczas na skutek otwarcia kanałów potasowych i obniżenia wewnątrzkomórkowego stężenia cAMP.
Hamowanie presynaptyczne
W zależności od tego z jaką częścią neuronu łączy się wypustka aksonalna wyróżniamy trzy rodzaje połączeń synaptycznych:
- akso-dendrytyczne : połączenie między aksonem i dendrytem;
- akso-somatyczne: połączenie między aksonem i ciałem neuronu;
- akso-aksonalne: połączenie między dwoma aksonami.
Zjawisko hamowania presynaptycznego zachodzi dzięki obecności połączeń aksonalno-aksonalnych. Polega ono na zahamowaniu przekazu impulsów nerwowych między komórkami nerwowymi, w skutek depolaryzacji błony presynaptycznej przez neurotransmitery, wydzielane przez inny akson, znajdujący się w sąsiedztwie hamowanej synapsy, tworzący synapsę akso-aksonalną.
Przewodzenie włóknami bezrdzennymi
Fizjologicznie źródłem pobudzenia dla początkowego odcinka aksonu jest potencjał pobudzający powstający w ciele komórki nerwowej. Jednak zarówno w organizmie, jak i po wyizolowaniu włókna nerwowego, można wywołać jego pobudzenie przez zadziałanie bodźcem na dowolny odcinek włókna nerwowego. Wówczas przepływ impulsu nerwowego zachodzi w obu kierunkach od miejsca zadziałania bodźca. Przewodzenie w kierunku ciała neuronu nazywamy przewodzeniem antydromowym. Natomiast przewodzenie w kierunku zakończenia aksonu to przewodzenie ortodromowe.
Postsynaptyczny potencjał pobudzający powoduje szybki napływ jonów sodowych do wnętrza komórki czego skutkiem jest depolaryzacja błony komórkowej. Napływ jonów sodowych jest jednak tak duży, że powoduje on szybkie chwilowe odwrócenie potencjału panującego wewnątrz komórki na dodatni, co opisywane jest jako nadstrzał - dodatni potencjał iglicowy.
Inaktywacja kanałów sodowych występująca, gdy potencjał iglicowy osiąga swoje maksymalne wartości, powoduje zahamowanie dalszego napływu jonów sodowych do wnętrza komórki. W tym samym czasie otwierają się kanały potasowe umożliwiając wypływ jonów potasu na zewnątrz komórki. W wyniku tych procesów dochodzi od repolaryzacji błony komórkowej. Ostatecznie równowaga jonów jest przywracana dzięki pracy pompy sodowo- potasowej, która zapewnia ponowną spoczynkową polaryzację błony komórkowej aksonu.
Przewodzenie włóknami rdzennymi
Pod wpływem działania bodźca dochodzi do nagłego napływu jonów sodowych do wnętrza aksonu, który powoduje depolaryzację jego początkowego odcinka. W przypadku włókien rdzeniowych depolaryzacja szerzy się dalej w sposób skokowy. W związku z przepływem jonów sodowych do depolaryzacji kolejnych odcinków włókna dochodzi w miejscu cieśni węzłowych, gdzie znajduje się największe nagromadzenie kanałów sodowych. Taki skokowy sposób przewodzenia impulsu nerwowego jest znacznie szybszy niż jego ciągłe przewodzenie we włóknach bezrdzennych.
Częstotliwość przewodzenia impulsów we włóknach nerwowych jest zmienna i często prowadzi to do powstania salw impulsów nerwowych. We włóknach nerwowych zaopatrujących komórki mięśniowe częstotliwość przewodzenia impulsów waha się od 60Hz do 80Hz. Natomiast, żeby wywołać maksymalny skurcz mięśnia częstotliwość docierających do niego impulsów nerwowych może osiągnąć nawet 150Hz.
Rodzaje włókien nerwowych
Ze względu na obecność osłonki mielinowej włókna nerwowe dzielimy na:
- włókna zmielinizowane- otoczone osłonka mielinową
- włókna niezmielinizowane- pozbawione osłonki mielinowej.
Ze względu na kierunek przewodzenia impulsów nerwowych włókna nerwowe dzielimy na:
- aferentne- wstępujące, przewodzące impulsy z obwodu dośrodkowo do ośrodka nerwowego
- eferentne- zstępujące, przewodzące impulsy nerwowe z ośrodka nerwowego odśrodkowo na obwód.
Różnice dotyczą również szybkości z jaką przewodzone są impulsy, czasu trwania potencjału iglicowego, długości czasu refrakcji (braku wrażliwości na bodźce), uwalnianych na zakończeniach aksonów neurotransmiterów i substancji modulujących.
Biorąc pod uwagę zarówno morfologiczne jak i czynnościowe cechy włókien nerwowych podzielono je na kilka grup:
- Grupa A- obejmuje czuciowe (aferentne) i somatyczne (eferentne) włókna zmielinizowane;
- Grupa B- obejmuje zmielinizowane włókna układu autonomicznego uwalniające na swoich zakończeniach acetylocholinę;
- Grupa Cs- obejmuje włókna niezmielinizowane uwalniające na swoich zakończeniach noradrenalinę;
- Grupa Cdr- obejmuje aferentne włókna niezmielinizowane, wnikające w korzeniach grzbietowych do rdzenia kręgowego.
Rolą włókien aferentnych grup A i Cdr jest przewodzenie impulsów nerwowych z receptorów do ośrodków nerwowych w układzie nerwowym. Natomiast włókna eferentne grup A i B wchodzą w skład nerwów czaszkowych i rdzeniowych, w których przewodzą impulsy z ośrodkowego układu nerwowego do narządów efektorowych.
