TECHNICAL FIELD
The present invention relates to a novel N-acetylglucosaminyl-transferase (GlCNAc transferase) which recognizes a specific sugar chain structure in a saccharide and introduces thereinto a GlcNAc β1→4 branching structure.
BACKGROUND ART
1. Glycoproteins
Most of proteins occurring in nature are not simple proteins composed of amino acids alone, but “mature” proteins having sugar chains and other substances such as phosphates and lipids attached thereto. Therefore, the development of simple protein-type products produced by Escherichia coli as a host has involved various problems because such products lack the maturing process of proteins. Since all of secretion-type physiologically active proteins (e.g. cytokines) are glycoproteins with a few exceptions, the function and the role of sugar chains have attracted attention as the most important point in the development of biological pharmaceuticals.
Sugar chains in glycoproteins are classified roughly into Asn-linked type, mucin-type, O-linked GlcNAc type, GPI anchor type and proteoglycan type [Makoto Takeuchi, “Glycobiology Series 5: Glycotechnology”, Kihata, Hakomori and Nagai (eds.), Kodansha Scientific Co., (1994), 191-208]. Each of these types of sugar chains has its own biosynthesis pathway and a discrete physiological function. Asn-linked sugar chains are distributed widely in molds, yeasts, insects, plants and animals. The basic biosynthesis pathway for Asn-linked sugar chains is conserved beyond species (FIG. 1). A sugar chain(s) characteristic of a specific species is(are) formed on the outer side (called the “non-reducing terminal side”) of the core sugar chain moiety which is common in the biosynthesis of Asn-linked sugar chains. A mannan-type sugar chain in whicha α 1,3- and α 1,2-branching mannose residues attach to a main chain extending via α 1,6 linkages is a sugar chain structure characteristic of fungi such as yeasts (see Panel a in FIG. 2) [Hiroshi Nakajima, Sugar Chain Technology, Industry Survey Association (1992), 384-397]. On the other hand, in insects, plants and animals., extension of mannose residues is not observed; instead, a high mannose type sugar chain is formed which is a sugar chain transferred from a dolichol intermediate and only trimmed (see Panel c in FIG. 2). A unique structure having characteristic xylose or the like (see Panel b in FIG. 2) is also observed in insects, plants and mollusks. In animals, characteristic sugar chain structures such as complex type sugar chain (Panel e in FIG. 2) and hybrid type sugar chain (Panel d in FIG. 2) are observed; in the former, GlcNAc branching structures are formed in a once trimmed sugar chain, and addition of other kinds of monosaccharides such as galactose and sialic acid forms complicated structures; in the latter, both a complex type sugar chain and a high mannose type sugar chain are present [Kiyoshi Furukawa, Sugar Chain Technology, Industry Survey Association (1992), 64-75].
Such sugar chains as described above are conferred on most of cell surface proteins and secretion proteins, and are thought to play important roles which determine the natures and properties of cells and proteins. Among all, the portion of a sugar chain structure which forms a branch elongating like antennas from the common core sugar chain is called a sugar chain branching structure. This structure is believed to have a function to give an organism recognition ligand (i.e., the end portion of the sugar chain) a high degree of freedom to thereby provide chances for multipoint recognition and another function to maximize the protection ability for the protein moiety by greatly increasing the space-occupying volume (Takeuchi et al., supra). Therefore, by controlling the branching structure of sugar chains, it is possible to modify the physiological functions, sush as itn vivo stability, in vivo kinetics and organ-targeting properties of glycoproteins in various ways. In view of this, technology to control branching structures of sugar chains is expected as biotechnology of the next generation for the development of glycoprotein-type pharmaceuticals which are “tender to humans”.
2. Physiological Significance of Glycoprotein Sugar Chains
Sugar chains of secretion type glycoproteins exhibit excellent functions in biosynthesis, intracellular sorting, masking of antigenicity, in vivo stability and organ-targeting properties of glycoproteins. Sugar chains of cell surface proteins are known to change in response to changes in cells (such as differentiation, change to a morbid state, canceration). In particular, it has been reported that there is a close relation between the metastasis of cancer and the branching structure of sugar chains.
(1) Masking of Antigenicity
It is considered that sugar chains have a high degree of freedom in terms of steric structure and thus are moving freely like propellers. Therefore, protein molecules such as proteases and antibodies against proteins not having affinity to sugar chains are shook off by the sugar chains and thus cannot gain access to the protein moiety. As a result, even if there is antigenicity in the peptide moiety near the sugar chain binding site, antibody molecules cannot have access to the peptide moiety. Thus, an antigen-antibody reaction is extremely difficult to occur. Further, when a glycoprotein has been captured by a macrophage and the degradation products are presented as antigen, receptors are difficult of access to the peptides around the sugar chain binding site. Thus, antigenic stimulation is difficult to occur. Actually, it is reported that when sugar chains have been introduced into the central portion of the antigenic peptide of ovalbumin lysozyme, the binding of MHC class II molecules to the antigen is remarkably inhibited [Mouritsen, S., Meldal, M., Christiansen-Brams, I., Elsner, H. and Werdelin, O., Eur. J. Immunol., (1994), 24, 1066-1072]. The effect of such masking of antigenicity becomes greater as the volume occupied by sugar chains is greater. Thus, it is considered that the development of a branching structure contributes to the effect of such masking greatly.
(2) In Vivo Stability
With respect to erythropoietin which is the first glycoprotein-type pharmaceutical ever produced from a transgenic animal cell as a host, the functions of sugar chains thereof have been studied thoroughly. As a result, it has been shown that the sugar chains of erythropoietin work inhibitorily against the binding of erythropoietin with its receptor but make a decisive contribution to the retaining of the active structure and the improvement of in vivo kinetics; as a whole, the sugar chains have been shown to be essential for expression of the pharmacological activity of erythropoietin (Takeuchi, M. and Kobata, A., Glycobiology (1991), 1, 337-346). In particular, a strong correlation between the number of antennae in sugar chains and the pharmacological effect of erythropoietin has been found, and thus the importance of its branching structure (a branching structure formed by GlcNAc residues attaching to the core sugar chain) which never attracted attention has been made clear for the first time [Takeuchi, M., Inoue, N., Strickland, T. W., Kobata, M., Wada, M., Shimizu, R., Hoshi, S., Kozutsumi, H., Takasaki, S. and Kobata, A., Proc. Natl. Acad. Sci. USA, (1989), 86, 7819-22]. The major cause of the above phenomenon is explained as follows: erythropoietin without developed branching structure is cleared rather rapidly in kidney and, as a result, the in vivo residence time of such erythropoietin becomes shorter [Misaizu, T., Matsuki, S., Strickland, T. W., Takeuchi, M., Kobata, A. and Takasaki, S., Blood, (1995), 86, 4097-4104].
(3) Organ Targeting Property
Most of biological tissues have lectin-like receptors and use then in cell-cell interactions or to uptake glycoproteins from blood. The asialoprotein-binding lectin in liver is a representative example of a clearance system for aged glycoproteins [Toshihiro Kawasaki, Sugar Chain Technology, Industry Survey Association (1992), 125-136]. In addition, selectin contained in vascular endothelial cells, platelets and leucocytes (Kawasaki, supra) and the lectin receptor present on the surface of macrophages and NK cells (Kawasaki, supra) are well known. Furthermore, not only glycoproteins but also cells are known to gather in a specific tissue using sugar chains as ligands. Cases of the homing of bone marrow cells [Tatsuo Irimura, “Glycobiology Series 3: Glycobiology in Cell Society”, Katsutaka Nagai, Senichiro Hakomori and Akira Kobata (eds.), Kodansha Scientific Co., (1993), 127-175] and the recruiting of neutrophiles to inflammatory sites (Irimura, supra) are examined in detail. Putting all these things together, it can be well assumed that glycoproteins and cells have, via their sugar chain structures, a targeting property toward specific organs or tissues presenting a lectin receptor in blood circulation, although such a targeting system is not found in all organ. This means that drug delivery by means of sugar chains is possible. In such drug delivery, the affinity of lectin for sugar chains is greatly influenced by the degree of freedom and the number of sugar chain ligands. Therefore, the branching structure of sugar chains will be the most important point in such drug delivery.
(4) Correlation between Cells' Change into Morbid State and Sugar Chain Branched Structure thereof [Junko Kato, Naoko Suzuki,” Sugar Chain Technology and Development of Pharmaceutical”, Foundation for the Relief and Study of Injury Caused by Pharmaceutical Side Effect (ed.), Yakugyo-Jiho-Sha, (1994), 107-13214]
Once a plant lectin called L-PHA was developed as a probe to detect a multi-branching type sugar chain structure, it has become possible to examine various morbid tissue samples. As a result, a tendency has been found that some types of cancer cells, particularly, cancer cells with a high metastasis ability are stained well with L-PHA. Thus, researchers have become aware of the correlation between the branching structure of sugar chains and the metastasis ability of cancer cells. Human chorionic gonadotropin (hCG) is a glycoprotein hormone vigorously biosynthesized in villus tissues at an early stage of pregnancy. Since a considerable amount of hCG is discharged into urine, hCG is clinically utilized as an indicator of pregnancy. The Asn-linked sugar chains mainly formed by the mono- and biantennary complex type chains are characteristic to hCG. As cancer increases its malignancy from trophoblastoma to invasive mole and from invasive mole to choriocarcinoma, it is reported that 2,4,2 type tri-antennary sugar chains and abnormal biantennary sugar chains (both are formed through the action of GnT-IV on normal biantennary and mono-antennary sugar chains, respectively) appear in the sugar chains of hCG [Katsuko Yamashita, Protein, Nucleic Acid and Enzyme (1992), 37, 1880-1888]. As a cause of this phenomenon, it is suggested that the activity of GnT-IV increases as the malignancy of choriocarcinoma progresses.
γ-Glutamyltranspeptidase (γ-GTP) is a glycoprotein occurring specifically abundant in liver. Since the serum γ-GTP level drastically increases when there is a liver disease, this level is used as clinical indicator of a liver disease. Further, Yamashita et al. [Yamashita, K., Totani, K., Iwaki, Y., Takamisawa, I., Takeishi, N., Higashi, T., Sakamoto, Y. and Kobata, A., J. Biochem., (1989), 105, 728-735] have found that, as a result of canceration of cells, the sugar chain structure of γ-GTP changes abnormally in its branching structure similar to those in abnormal hCG; thus, they have reported the correlation between canceration and the activation of GnT-IV. The Asn-linked sugar chains of γ-GTP derived from healthy human liver cells are composed mainly of the biantennary complex type sugar chain with small amounts of tri-antennary and tetra-antennary sugar chains mixed therein. In contrast, a remarkable increase in the degree of branching was observed in the Asn-linked sugar chains of γ-GTP derived from human hepatoma cells. At the same time, though small in amounts, high mannose type sugar chains and abnormal biantennary sugar chains (both of which were not observed in γ-GTP from normal cells) appeared. As a cause of these changes in sugar chain structure, a possibility is suggested that N-acetylglucosaminyltransferase IV (GnT-IV) and V (GnT-V) are activated in relation to canceration of liver cells (Yamashita et al., supra).
It is also reported that the sugar chain branching structure of a glycoprotein in cells is greatly changed by viral infection (Yamashita et al., supra). BHK cells have sugar chain structures with branching up to tetraantennary type. When BHK cells are transformed with polyomavirus, biantennary type sugar chains decrease in the glycoprotein sugar chains produced by the cells, while tetraantennary type sugar chains and the N-acetyllactosamine repeat structures increase; as a whole, a remarkable increase in the number of branches was recognized [Takasaki S., Ikehira, H. and Kobata A., Biochem. Biophys. Res. Commun., (1980), 90, (3), 735-742]. As a cause of the above change, activation of GnT-IV, GnT-V and i-GnT may be considered.
3. Enzymes relating to the Sugar Chain Branching Structures of Glycoproteins
The complex type sugar chain which is a glycoprotein sugar chain structure characteristic of animals has a complicated branching structure in which N-acetylglucosamine (GlcNAc) residues are attaching to the common core structure in various manners (Kiyoshi Furukawa, supra) (FIG.
1). Since this branching structure is closely related to in vivio and in vivo stability, localization, biological activity and pharmacological property of glycoproteins (Makoto Takeuchi, supra), the process of biosynthesis of the branching structure has been investigated in detail. By using inventive substrates H. Schachter et al. have discriminated the various enzyme activities in hen oviduct to thereby predict the presence of GlcNAc branch forming enzymes from GnT-I to GnT-VI (group of GlcNAc glycosyltransferases; FIG. 3) [Glesson, P. A. and Schachter, H., J. Biol. Chem., (1983), 258, 6162-6173]. Thereafter, GnT-I [Kumar, R., Yang, J., Larsen, R. D. and Stanley P., Proc. Natl. Acad. Sci. USA, (1990), 87, 9948-9952; Sarkar, M., Hull, E., Nishikawa, Y., Simpson, R. J., Moritz, R. L., Dunn, R. and Schachter, H., Proc. Natl. Acad. Sci., USA, (1991), 88, 234-238], GnT-II [D'Agostaro, G A., Zingoni, A., Moritz, R L., Simpson, R J., Schachter, H. and Bendiak, B., J. Biol. Chem., (1995), 270, 15211-21], GnT-III [Nishikawa, A., Ihara, Y., Hatakeyama, M., Kangawa, K. and Taniguchi, N., J. Biol. Chem., (1992), 267, 18199-18204] and GnT-V [Shorebah, M. G., Hindsgaul, O . and Pierce, M., J. Biol. Chem., (1992), 267, 2920-2927; Gu, J., Nishikawa, A., Turuoka, N., Ono, M., Yamaguchi, N., Kangawa, K. and Taniguchi, N., J. Biochem., (1993), 113, 614-619] were successively purified, and the genes thereof were cloned. However, with these known GlcNAc transferases alone, it is impossible to form the main sugar chain (tetraantennary type; see the formula below) found in
α 1 acid glycoprotein known as a representative human blood glycoprotein [Yoshima, K., Tsuji, T., Irimura, T. and Osawa, T, J. Biol. Chem., (1984), 256, 10834-10840] and erythropoietin [Takeuchi, M., Takasaki, S., Shimada, M. and Kobata, A., J. Biol. Chem., (1990), 265, 12127-12130]. Therefore, an N-acetylglucosaminyltransferase having such substrate specificity and reaction specificity that are expected in GnT-IV has been searched for as a missing link.
In addition to those mentioned above, the following N-acetyl-glucosaminyltransferases have been purified or the genes thereof have been cloned: a transferase acting on mucin-type sugar chains [Bierhuizen, M. F., Maemura, K. and Fukuda, M., J. Biol. Chem., (1994), 269, 4473-4479], a transferase acting on glycolipids, and a transferase forming the sugar chain epitope known as I·i antigenic structure [Kawashima, H., Yamamoto, K., Osawa, T. and Irimura, T., J. Biol. Chem., (1993), 268, 27118-27126; Bierhuizen, M. F., Mattei, M. G. and Fukuda, M., Genes Dev., (1993), 7, 468-478]. However, the substrate specificity of these transferases and the mode of binding of the GlcNAc group transferred by these transferases are different from those of GnT-IV. Any of these transferases does not yield products resembling GnT-IV products.
DISCLOSURE OF THE INVENTION
It is an object of the present invention to provide an enzyme having β1→4 N-acetylglucosaminyltransferase (hereinafter referred to as “GnT-IV”) activity; a gene encoding the enzyme; a recombinant DNA comprising the gene; a cell containing the recombinant DNA; a method for producing an enzyme protein having GnT-IV activity comprising culturing the cell in a medium; and a saccharide in which the sugar chains are modified with GnT-IV.
Toward the solution of the above assignments, the present inventors have made intensive and extensive researches. As a result, the inventors have isolated and purified a GnT-IV enzyme protein from bovine small intestine, and characterized the biochemical properties of the protein. Then, the inventors have succeeded in cloning a gene coding for bovine GnT-IVa from a cDNA library and mRNA from the small intestine based on a partial amino acid sequence of the above enzyme protein. Further, based on bovine GnT-IVa gene, the inventors have succeeded in cloning two genes coding for human GnT-IVa and human GnT-IVb, from cDNA libraries and mRNAs from human liver and human lung, respectively. The present invention has been completed by confirming that the products of these genes exhibit GnT-IV activity.
The first invention of the present application relates to a GnT-IV having an activity to produce a saccharide having a partial structure represented by the formula below:
using UDP-GlcNAc as a sugar donor and a saccharide having a partial structure represented by the formula below as a sugar receptor:
The second invention relates to a GnT-IV consisting of the amino acid sequence shown in SEQ ID NO: 18 or the amino acid sequence shown in SEQ ID NO: 18 which has addition, deletion or substitution of one or more amino acid residues and yet which produces GnT-IV activity; a GnT-IV consisting of the amino acid sequence shown in SEQ ID NO: 24 or the amino acid sequence shown in SEQ ID NO: 24 which has addition, deletion or substitution of one or more amino acid residues and yet which produces GnT-IV activity; and a GnT-IV consisting of the amino acid sequence shown in SEQ ID NO: 37 or the amino acid sequence shown in SEQ ID NO: 37 which has addition, deletion or substitution of one or more amino acid residues and yet which produces GnT-IV activity.
The third invention relates to a GnT-IV gene coding for a GnT-IV consisting of the amino acid sequence shown in SEQ ID NO: 18 or the amino acid sequence shown in SEQ ID NO: 18 which has addition, deletion or substitution of one or more amino acid residues and yet which produces GnT-IV activity; a GnT-IV gene coding for a GnT-IV consisting of the amino acid sequence shown in SEQ ID NO: 24 or the amino acid sequence shown in SEQ ID NO: 24 which has addition, deletion or substitution of one or more amino acid residues and yet which produces GnT-IV activity; a GnT-IV gene coding for a GnT-IV consisting of the amino acid sequence shown in SEQ ID NO: 37 or the amino acid sequence shown in SEQ ID NO: 37 which has addition, deletion or substitution of one or more amino acid residues and yet which produces GnT-IV activity; a GnT-IV gene consisting of the nucleotide sequence shown in SEQ ID NO: 17; a GnT-IV gene consisting of the nucleotide sequence shown in SEQ ID NO: 23; and a GnT-IV gene consisting of the nucleotide sequence shown in SEQ ID NO: 36.
The fourth invention relates to a recombinant DNA obtainable by inserting any of the above GnT-IV gene into a vector DNA; and a chromosomal fragment comprising a part or all of any one of the above GnT-IV gene.
The fifth invention relates to a host cell carrying the above recombinant DNA; and a host cell into which the above chromosomal fragment is artificially introduced.
The sixth invention relates to a method for producing a GnT-IV comprising culturing the above host cell in a medium and recovering the GnT-IV from the resultant culture; and a method for producing a GnT-IV comprising recovering the GnT-IV enzyme from the secreta, body fluids or homogenete originated from the above host cell.
The seventh invention relates to a method for purifying the GnT-IV from biological samples.
The eighth invention relates to a saccharide of which the sugar chain structure is modified with the GnT-IV.
Hereinbelow, the present invention will be described in detail.
The GnT-IV gene of the invention can be isolated as described below.
Isolation of Bovine GnT-IVa Gene
First, a microsome fraction from bovine small intestine solubilized with a detergent is subjected to a series of purification procedures using anion exchange chromatography, copper chelate chromatography, two-step affinity chromatography using a substrate analogue and gel filtration to thereby obtain a purified sample of GnT-IV enzyme. The resultant purified sample is subjected to SDS-PAGE and then transferred onto a PVDF membrane. The transferred protein, as it is or after restricted hydrolysis, is analyzed with a gas phase amino acid sequencer to obtain a partial amino acid sequence for the GnT-IV enzyme.
Subsequently, an RT-PCR is performed on the RNA extracted from the animal cells (i.e., bovine small intestine) as a template using primers designed based on the partial amino acid sequences determined above. Further, using a fragment obtained by the RT-PCR as a probe, the GnT-IV gene of interest is screened from a cDNA library from the above-mentioned tissue by plaque hybridization. A cDNA fragment contained in the resultant positive plaque is cut out and subcloned into a vector such as pUC19, followed by analysis of the nucleotide sequence thereof. If the full length of the gene coding for the protein of interest is not contained in the fragment, plaque hybridization is performed again using a part of the subcloned cDNA fragment as a probe. Alternatively, terminal portions of the cDNA of interest are obtained by RACE or the like based on the information on the nucleotide sequence obtained above. The thus obtained GnT-IV gene (which is named GnT-IVa aferward) is subjected to analysis of its entire nucleotide sequence. Subsequently, the amino acid sequence is translated from the gene having the above-mentioned nucleotide sequence. This amino acid sequence is as shown in SEQ ID NO: 18.
Isolation of Human GnT-IVa and GnT-IVb Genes
Human GnT-IVa and GnT-IVb genes can be obtained by performing a RT-PCR using RNA extracted from a human tissue (liver or lung) and based on the information on the nucleotide sequence of bovine GnT-IVa gene as obtained above, followed by screening of a cDNA library from the above tissue. The resultant human GnT-IVa and GnT-IVb genes are subjected to analysis of their entire nucleotide sequences. Subsequently, the amino acid sequences are translated by these genes. These amino acid sequences are as shown in SEQ ID NOS: 24 and 37.
In order to obtain a DNA coding for the amino acid sequence shown in SEQ ID NO: 18, 24 or 37 having addition, deletion or substitution of one or more amino acid residues, a number of methods may be used. For example, a method of treating DNA with a mutagen to induce point mutation or a deletion mutation; a method comprising cleaving DNA selectively, removing or adding a selected nucleotide and then ligating DNA; site-specific mutagenesis; and the like may be enumerated.
The GnT-IV protein of the invention can be produced by preparing a recombinant vector into which a DNA coding for the GnT-IV of the invention obtained by the method described above is inserted downstream of a promoter, introducing the vector into a host cell and culturing the resultant cell. The vector DNA used for this purpose may be either plasmid DNA or bacteriophage DNA. For example, pSVL vector (Pharmacia, Sweden) shown in an Example described later may be used. As the host cell into which the resultant recombinant DNA is introduced, any cell that is conventionally used in recombinant DNA techniques may be used, for example, a prokaryotic cell, an animal cell, a yeart, a fungi, an insect cell. Specific examples include Escherichia coli as a prokaryotic cell and CHO cells from chinese hamster or COS cells from monkey as an animal cell.
The transformation of the host cell described above is performed by conventional methods for each host. For example, if the host is E. coli, a vector comprising the recombinant DNA is introduced by the heat shock method or electroporation into competent cells prepared by the calcium method or the like. If the host is yeast, a vector comprising the recombinant DNA is introduced by the heat shock method or electroporation into competent cells prepared by the lithium method or the like. If the host is an animal cell, a vector comprising the recombinant DNA is introduced into the cell at the growth phase or the like by the calcium phosphate method, lipofection or electroporation.
By culturing the thus obtained transformant in a medium, the GnT-IV protein is produced.
In the cultivation of a transformant, any medium may be used as long as the host is viable in it. For example, LB medium or the like may be used if the host is E. coli. If the host is yeast, YPD medium or the like may be used. If the host is an animal cell, Dulbecco's medium supplemented with an animal serum or the like may be used. The cultivation is performed under conditions conventionally used for the host. For example, if the host is E. coli, cells are cultured at about 30-37 ° C. for about 3-24 hours with, if necessary, aeration and/or agitation. If the host is yeast, cells are cultured at about 25-37 ° C. for about 12 hours to 2 weeks with, if necessary, aeration and/or agitation. If the host is an animal cell, cultivation is performed at about 32-37 ° C. under 5% CO2 and 100% humidity for about 24 hours to 2 weeks with, if necessary, change of the aeration conditions and/or agitation.
After the cultivation, the cultured microorganism or cells are disrupted using a homogenizer, French press, sonication, lysozyme and/or freeze-thawing to thereby elute the GnT-IV protein outside the microorganism or cells. Then, the protein can be obtained from soluble fractions. If the protein of interest is contained in insoluble fractions, the insoluble fractions are collected by centrifugation after disruption of the microorganism or cells. Then, the protein may be solubilized with a buffer containing guanidine hydrochloride or the like for recovery. Alternatively, the cultured microorganism or cells may be disrupted directly with a buffer containing a protein denaturing agent such as guanidine hydrochloride to thereby elute the protein of interest outside the microorganism or cells.
Purification of the GnT-IV protein from the above supernatant may be performed by the method described in Example 1. Alternatively, this purification may be performed by appropriately combining conventional separation/purification methods. These conventional separation/purification methods include, but are not limited to, centrifugation, salting out, solvent precipitation, dialysis, ultrafiltration, partition chromatography, gel filtration, capillary electrophoresis, TLC, ion exchange chromatography, metal chelate chromatography, affinity chromatography, reversed phase chromatography and isoelectric focusing.
The biochemical properties of the GnT-IV enzyme protein obtained from bovine small intestine as described above are as follows.
(1) Action
This enzyme protein produces a saccharide having a partial structure represented by the formula below:
using UDP-GlcNAc as a sugar donor and a saccharide having a partial structure represented by the formula below as a sugar receptor:
The saccharide as a sugar receptor means an oligosaccharide, polysaccharide, glycoconjugate (glycopeptide, glycoprotein, glycolipid or proteoglycan) or a derivative thereof.
(2) Substrate Specificity
When the sugar receptor is an oligosaccharide (for the structures of oligosaccharides, see FIG. 4), the enzyme protein exhibits reactivities of 0% toward core type oligosaccharides, 54% toward GnT-I product type oligosaccharides and 164% toward GnT-V product type oligosaccharides, wherein the reactivity of the enzyme protein toward GnT-II product type oligosaccharides is regarded as 100%.
The enzyme protein exhibits a reactivity of 46% toward a structure of GnT-II product type oligosaccharides in which fucose is attached via α1→6 linkage to the GlcNAc at the reducing terminus.
The enzyme protein exhibits a reactivity of 0% toward a structure of GnT-II product type oligosaccharides in which the GlcNAc on the α1→3 mannose is lacking.
The enzyme protein exhibits a reactivity of 16% toward a structure of GnT-II product type oligosaccharides in which galactose is attached viag β1→4 linkage to the GlcNAc on the α 1→6 mannose, and a reactivity of 0% toward a structure of GnT-II product type oligosaccharides in which galactose is attached via β1 4 linkage to the GlcNAc on the α1→3 mannose.
The enzyme protein exhibits a reactivity of 0% toward a structure of GnT-II product type oligosaccharides in which GlcNAc is attached via β 1→4 linkage to the β1→4 mannose.
(3) Molecular Weight
About 66 K as determined by SDS-PAGE (under non-reducing conditions). About 60 K after treatment with peptide N-glycosidase F. Since a shift of band is observed when peptide N-glycanase is used, the enzyme protein is thought to be a glycoprotein.
The apparent molecular weight as determined by filtration with a gel containing Triton X-100 is 77 K. Thus, it is thought that GnT-IV does not have a subunit structure and functions as a monomer.
The protein moiety of this enzyme deduced from the nucleotide sequence thereof consists of 535 amino acid residues and has a molecular weight of 61614.
(4) Optimum pH
The optimum pH for reaction is about 5.5. More than 50% of the maximum activity is observed in the range from pH 6.5 to 8.0.
(5) Inhibition, Activation and Stabilization
(i) Inhibition
The activity of this enzyme is inhibited by addition of 20 mM EDTA.
This enzyme is inhibited by UDP derivatives. The intensity of inhibition is in the following order: UDP>>UDP-Glc>UDP-GalNAc>>2′-deoxy UDP>UDP-hexanolamine>>UDP-Gal>UTP>UDP-glucuronic acid>UMP.
Uridine, TDP and CDP do not have inhibitory effect.
(ii) Activation
A divalent cation is essential for expression of the activity. Among divalent cations, Mn2+ shows the greatest effect. At a concentration of 7.5 mM, the respective effects of Co2+and Mg2+ are about 70% of that of Mn2+, and the effect of Ca2+ is about 10% of that of Mn2+. The effect of Mn2+ is greatest in the range from 5 to 20 mM.
(iii) Stabilization
Stabilizing effect is recognized in BSA and glycerol.
(6) Kinetic Parameters
When the saccharide as a receptor is an oligosaccharide (for the structures of oligosaccharide, see FIG. 4):
(i) under assay conditions in which the enzyme is reacted in 50 μl of 125 mM MOPS buffer (pH 7.3) containing 0.8 mM receptor substrate, 20 mM UDP-GlcNAc, 7.5 mM MnCl2, 200 mM GlcNAc, 0.5% (w/v) Triton X-100, 10% glycerol and 1% BSA at 37° C. for 4 hours:
Km and Vmax values toward GnT-II product type oligosaccharide are 0.73 mM and 3.23 μM/min, respectively.
Km and Vmax values toward GnT-V product type oligosaccharide are 0.13 mM and 1.75μM/min, respectively.
When GnT-II product type oligosaccharide is the receptor substrate, Km value toward UDP-GlcNAc is 0.22 mM.
(ii) under assay conditions in which the enzyme is reacted in 125 mM MOPS buffer (pH 7.3) containing 120 mM UDP-GlcNAc, 7.5 nM MnCl2, 0.5% (w/v) Triton X-100, 10% glycerol and 1% BSA at 37° C. for 4 hours:
Km and Vmax values toward GnT-II product type oligosaccharide are 0.59 mM and 0.74 mM/min/mg, respectively.
Km and Vmax values toward GnT-V product type oligosaccharide are 0.14 mM and 0.47 mM/min/mg, respectively.
(7) GnT-IV Family
The homology between bovine GnT-IVa and human GnT-IVa is 91% at the nucleic acid level and 96% at the amino acid level.
All of the partial amino acid structures contained in the purified GnT-IV from bovine small intestine are encoded in the bovine GnT-IVa gene.
Human GnT-IVb and human GnT-IVa have 63% homology at the nucleic acid level and 62% homology at the amino acid level. However, they are entirely different in the C-terminal and N-terminal regions.
From the biochemical properties described above, the GnT-IV of the invention has been recognized as a novel enzyme in the point that this enzyme is able to perform the following reaction which conventional enzymes cannot perform:
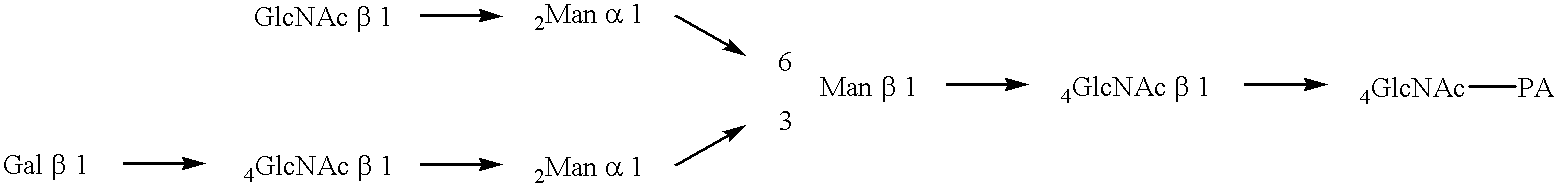
FIG. 1 shows the biosynthetic pathway of Asn-linked sugar chains.
FIG. 2 shows variations of the Asn-linked sugar chain (revised from FIG. 1 in Makoto Takeuchi, Wako Purechemical Newsletter 64, 18-19, 1996).
a. Mannan type: a sugar chain structure characteristic of fungi such as yeasts and molds.
b. Xylo-high-mannose type: a structure characteristic of plants, mollusks and insects.
c. High mannose type: a structure commonly seen in plants, insects and animals.
d. Hybrid type: a structure commonly seen in insects and animals.
e. Complex type: a structure characteristic of animals.
f. Prokaryotic cells: have no system for biosynthesis of Asn-linked sugar chains.
The portion boxed with dotted lines represents the common core sugar chain.
FIG. 3 shows the positions of GlcNAc transfer by various GlcNAc transferases (GlcNAc glycosyltransferases).
FIG. 4 shows the designations and structures of oligosaccharides.
FIG. 5 shows a high performance liquid chromatogram for GnT-IV reaction products.
FIG. 6 shows the results of analysis of GnT-IV by Q-Sepharose FF chromatography.
FIG. 7 show the results of analysis of GnT-IV by copper chelate Sepharose FF chromatography.
FIG. 8 shows the results of analysis of GnT-IV by UDP-Hexanolamine Agarose affinity chromatography (I).
FIG. 9 shows the results of analysis of GnT-IV by UDP-Hexanolamine Agarose affinity chromatography (II).
FIG. 10 shows the results of analysis of GnT-IV by Superdex 200 gel chromatography.
FIG. 11 is a photograph showing the results of SDS-PAGE (SDS polyacrylamide gel electrophoresis) of purified GnT-IV.
FIG. 12 shows the results of native gel electrophoresis of purified GnT-IV and the activity thereof.
FIG. 13 shows Smith degradation profile of GnT-IV, -V and VI product type oligosaccharides.
FIG. 14 shows the results of 1H-NMR (30° C.) of the GnT-IV reaction product.
FIG. 15 shows the optimum pH for GnT-IV.
FIG. 16 shows the optimum Mn2+ concentration for GnT-IV.
FIG. 17 shows the results of analysis by SDS-PAGE and fluorochromatography of glycoproteins which are the reaction products of GnT-IV.
Lanes 1 and 2: 7.6 μg of asialo agalacto human transferrin
Lane 3: 7.6 μg of asialo human transferrin
Lanes 4 and 5: 2.8 μg of asialo agalacto, CHO cell-derived recombinant human erythropoietin
Lanes 6 and 7: 1.3 μg of asialo agalacto fetuin
Lanes 1, 4 and 6 represent mock experiments in which reaction was performed without GnT-IV. M represents molecular markers (Bio-Rad). PM represents pre-stained molecular markers (Bio-Rad, USA).
GnT-IV reaction conditions: To 10 μl of a solution containing 0.702 mnol/hr of GnT-IV, a substrate glycoprotein equivalent to 1.6 nmol of biantennary type sugar chains (for fetuin alone, the sugar chain content was 1.6 nmol) and 450 nCi of UDP-[14C] GlcNAc, an equal volume of an assay mixture (250 mM MOPS buffer, pH 7.3, 400 mM GlcNAc, 20% glycerol, 1.0% (w/v) Triton X-100, 15 mM MnCl2, 1 mM UDP-GlcNAc) was added to obtain a reaction solution, which was incubated at 37° C. for 20 hours. One tenth of the resultant solution was analyzed by SDS-PAGE and fluorography.
For SDS-PAGE, 10-20% gradient gel (Daiichi Kagaku) was used. For fluorography, Amplify (Amersham) was used to expose the samples to X ray film for 20 hours.
FIG. 18 shows the open reading frame of human GnT-IVa and the region contained in pCore-His expression vector.
FIG. 19 shows the results of isoelectric focusing and Western analysis of erythropoietins produced by individual cell clones. Using two erythropoietin-producing strains and the same strains into which bovine and human GnT-IVa genes were introduced, respectively, the erythropoietin secreted by each strain was analyzed by isoelectric focusing and Western blotting using anti-erythropoietin antibody. On the left side, the positions of pI markers are shown.
BEST MODES FOR CARRYING OUT THE INVENTION
Hereinbelow, the present invention will be described in more detail with reference to the following Examples. However, the present invention is not limited to these Examples.
REFERENCE EXAMPLE 1
(1) Reagents Used in the Examples
Unless indicated otherwise, the reagents used were the highest grade products manufactured by Wako Purechemical Industries, Ltd.
(i) Pyridylaminated Oligosaccharides
Each of the pyridylaminated oligosaccharides used was obtained as described below. First, pyridylaminated oligosaccharides were prepared from human transferrin (apo type; Sigma, USA) according to the method of Tokugawa et al. [Biehuizen, M. F., Mattei, M. G. and Fukuda, M. (1993) Genes Dev., 7, 468-478]. The resultant material was treated with one or a combination of the following enzymes: Arethrobacter ureafaciens-derived sialidase (Nacalai Tesque), Asperugillus sp.-derived β-galactosidase (Toyobo), Jack bean-derived β-N-acetylhexosaminidase (Seikagaku Corp.), GnT-V active fraction in CHO-K1 cell extract (supernatant obtained by sonication of CHO-K1 cells in 2 volumes of 5 mM Tris-HCl buffer, pH 7.5, containing 2 mM MgCl2 and 1 mM PMSF, and then centrifugation at 900×g for 10 min), and the GnT-V active fraction in the solubilized fraction from bovine small intestine homogenate (for the method of preparation, see “Preparation of the Microsome Fraction” and “Solubilization” in Example 1). A part of the pyridylaminated oligosaccharides were prepared by treating PA-Sugar Chain 021 and 022 (Takara Shuzo) with the above enzymes. In both cases, the oligosaccharides prepared were purified by reversed phase chromatography using an ODS column (10×250 mm; Vydac, USA) before use.
(ii) Glycoprotein Substrates
Bovine fetuin (Sigma, USA) and CHO cell-derived recombinant human erythropoietin (Kirin Brewery) were subjected to the following pretreatment to purify them into relatively uniform glycoform. Briefly, 40-100 mg of the glycoprotein was applied to a ConA-Sepharose column (5 ml; Pharmacia, Sweden) equilibrated with 10 mM Tris-HCl buffer, pH 7.4, containing 1 mM MgCl2, 1 mM CaCl2 and 0.15 M NaCl to thereby obtain a glycoform with a low biantennary sugar chain content as the non-adsorbed fraction. Thereafter, the column was eluted with the above buffer containing 1.0 M α-methyl mannoside (Nacalai Tesque) to thereby obtain the fraction adsorbing a glycoform with a high biantennary sugar chain content. Thus, fetuin with a low biantennary sugar chain content and erythropoietin with a high biantennary sugar chain content were obtained. With respect to human transferrin, there is no need to purify this glycoprotein since almost all sugar chains thereof are biantennary.
The thus obtained fetuin and human transferrin were individually reacted with 1 U of sialidase and 0 or 107 U of β-galactosidase in 1 ml of 0.4 M sodium acetate buffer, pH 5.0, containing 4 mM MgCl2 at 37° C. for 16 hours to thereby obtain asialo or asialo agalacto glycoproteins. The erythropoietin with a high biantennary sugar chain content was reacted with 0.5 U of sialidase and 5 U of β-galactosidase in the same manner as described above to obtain an asialo agalacto glycoprotein.
Each of the thus obtained glycoprotein substrates was dialyzed against 50 mM ammonium acetate buffer, pH 7.3. Then, the amount of protein was determined with BCA protein assay (Pierce, USA) using BSA (bovine serum albumin) as a standard. Further, the protein was analyzed by SDS-PAGE (sodium dodesyl sulfate polyacrylamide gel electrophoresis). The thus prepared glycoproteins were used in the Examples.
(iii) RT-PCR (Reverse Transcription-Polymerase Chain Reaction)
For RT-PCR, Access RT-PCR System (Promega, USA) was used. For the amplification of a fragment of a gene of interest, Pfu polymerase (Stratagene, USA) was used.
(2) Equipment used in the Examples
(i) Gene Sequencing
ABI PLISM 377 DNA Sequencer (Perkin-Elmer, USA) was used.
REFERENCE EXAMPLE 2
Specific Assay for GnT-IV Activity
Generally, there are two methods for assaying GnT-IV activity: a method in which the transfer of radiolabeled GlcNAc to an oligosaccharide substrate is examined and a method in which the transfer of GlcNAc to a labeled oligosaccharide substrate is fractionally analyzed by HPLC or the like. Taniguchi et al. developed a method in which GnT-III, -IV and -V activities are simultaneously determined using the GnT-II product type oligosaccharide as a receptor [Nishikawa, A., Fujii, S., Sugiyama, T. and Taniguchi, N. (1988) Anal. Biochem., 170, 349-354]. However, this assay method as it is was inappropriate for the assay during purification of GnT-IV because the relative activity of GnT-IV is much lower than those of GnT-III and -V.
Then, the present inventors have developed a method for determining GnT-IV activity quantitatively and sensitively by increasing the amount of the acceptor pyridylaminated oligosaccharide to 10-fold compared to the amount used in the previous assay system [Tokugawa, K., Oguri, S. and Takeuchi, M. (1996) Glycoconjugate J., 13, 53-56]. Generally, it is very difficult to prepare such a large amount of acceptor oligosaccharides. However, according to the method of Tokugawa et al. [Tokugawa, K., Oguri, S. and Takeuchi, M. (1996) Glycoconjugate J., 13, 53-56], such oligosacchardes are readily prepared.
In Examples of the present invention, GnT-IV activity was assayed as described below.
The enzyme was reacted in 125 mM MOPS [3-(N-morpholino)propane-sulfonic acid] buffer, pH 7.3, containing 0.8 mM pyridylaminated oligosaccharide substrate (GnT-II product type oligosaccharide substrate), 20 mM UDP-GlcNAc, 7.5 mM MnCl2, 200 mM GlcNAc, 0.5%(w/v) Triton X-100, 10% glycerol and 1% BSA at 37° C. for 4 hours. Then, the reaction was terminated by boiling the solution for 2 minutes. After removal of solids with a 0.45 nm filter, 5 μ1 of the filtrate was analyzed with an ODS-80TM column (4.6×150 mm; TOSO) (FIG. 5) at 50° C. with 50 mM ammonium acetate buffer, pH 4.0, containing 0.15% (w/v) n-butanol at a flow rate of 1.2 ml/min. The fluorescence of pyridylamino groups was detected using excitation at 320 nm and emission at 400 nm.
EXAMPLE 1
Isolation and Purification of the Enzyme
(1) Screening of a Source of the Enzyme
A source of the GnT-IV enzyme to be purified was searched for by utilizing the assay method described above. It was found that the relative activity of GnT-IV to those of GnT-III and GnT-V in bovine small intestine is rather higher than the relative activities of GnT-IV in any other tissues as shown in Table 1. Thus, bovine small intestine was selected as a starting material for purification.
| TABLE 1 |
| |
| Search for Sources of GnT-IV Enzyme |
| |
Specific Activity (pmol/h · mg-protein) |
| Source of the Enzyme |
IV |
III |
V |
| |
| Cultured cells |
CHO |
10.8 |
0 |
1097 |
| |
Bowes |
12.0 |
341 |
150 |
| |
AH661) |
2.0 |
634 |
30 |
| |
Solid AH1) |
27 |
116 |
80 |
| |
Yoshida |
1.3 |
70 |
109 |
| |
sarcoma1) |
| Rat organs1) |
Small intestine |
17 |
280 |
68 |
| |
Heart |
9.4 |
11 |
10 |
| |
Spleen |
20 |
100 |
21 |
| |
Kidney |
1.9 |
1840 |
30 |
| |
Brain |
3.7 |
660 |
38 |
| Human1) |
Liver |
2.8 |
8.1 |
8.2 |
| Bovine organs |
Small intestine |
25 |
174 |
41 |
| |
Heart |
N.D. |
N.D. |
N.D. |
| |
Spleen |
10.9 |
0.7 |
10.9 |
| Colostrum |
|
N.D. |
N.D. |
N.D. |
| |
| N.D. below detection limit |
| 1)data from Nishikawa, A. et al., BBA 1035, 313-318 (1990) addition of EDTA (ethylenediamine tetra-acetic acid). A divalent cation is essential for its activity. Among divalent cations, Mn2+ shows the greatest effect, followed by Co2+ and Mg2+. Weak effect is recognized in Ca2+ and Fe2+. The optimum concentration of Mn2+ is around 10 mM as shown in FIG. 16. |
(2) Purification
Unless otherwise indicated, all the operations were performed at 4° C.
(i) Preparation of the Microsome Fraction
Two kilograms of bovine small intestine (obtained from a meat processor) was minced. Then, 4 volumes of an extraction buffer (10 mM Tris-HCl buffer, pH7.4, containing 0.25 M sucrose, 1 mM phenylmethylsulfonyl fluoride, 1 mM benzamidine hydrochloride, 1 mM dithiothreitol and 10 mg/ml antipain) was added thereto and homogenized with Polytron (Kinematica, Sweden). The resultant homogenate was centrifuged at 900×g for 10 minutes. Then, the supernatant was centrifuged further at 105,000×g for 60 minutes to thereby obtain the microsome fraction as a precipitate (Sample 1).
(ii) Solubilization
Sample 1 was suspended in 3 volumes of a solubilization buffer prepared by adding Triton-100 to the extraction buffer to give a final concentration of 1%. The supernatant was obtained by the centrifugation at 105,000×g for 60 minutes. The pellet was suspended again and collect the supernatant. The first and second extracts were combined (Sample 2).
(iii) Q-Sepharose FF Chromatography
Sample 2 was applied to Q-Sepharose FF Column (5×30 cm; Pharmacia, Sweden) pre-equilibrated with operation buffer 1 (20 mM Tris-HCl, pH 7.4, containing 1 mM benzamidine hydrochloride, 0.1% Triton X-100 and 20% glycerol) and then eluted by a linear gradient of 0-0.5 M KCl FIG. 6)(Sample 3).
(iv) Copper Chelate Sepharose FF Chromatography
Sample 3 was applied to Copper Chelate Sepharose FF Column (5×10 cm; Pharmacia, Sweden) pre-equilibrated with operation buffer 2 (obtainable by adding KCl to operation buffer 1 at a final concentration of 0.15 M). Then, non-adsorbed fractions were washed out with 5 volumes of operation buffer 2. Thereafter, the adsorbate was eluted by a linear gradient of 0.01 M glycine (FIG. 7). The resultant GnT-IV active fraction was pooled and concentrated with YM30 ultrafiltration membrane (Amicon, USA) (Sample 4).
(v) UDP-Hexanolamine Agarose Affinity Chromatography I
To UDP-Hexanolamine Agarose Affinity Column (1.2×4.5 cm; Sigma, USA) pre-equilibrated with operation buffer 3 (20 mM Tris-HCl, pH 8.0, containing 0.15 M KCl, 10 mM MnCl2, 0.05% Triton X-100 and 20% glycerol), one half of Sample 4 dialyzed against 1 mM benzamidine hydrochloride-added operation buffer 3 was applied. Then, non-adsorbed fractions were washed out with operation buffer 4 (20 mM Tris-HCl, pH 8.0, containing 10 mM MnCl2, 0.05% Triton X-100 and 20% glycerol). Thereafter, the adsorbate was eluted with operation buffer 4 to which 1 M (final concentration) KCl had been added (FIG. 8). The GnT-IV active fraction was pooled and dialyzed against operation buffer 5 (having the same composition as that of operation buffer 4 but having a pH of 7.4) (Sample 5).
(vi) UDP-Hexanolamine Agarose Affinity Chromatography II
Sample 5 was applied to UDP-Hexanolamine Agarose Affinity Column (1.0×6.5 cm; Sigma, USA) pre-equilibrated with operation buffer 5. Then, non-adsorbed fractions were washed out with operation buffer 5. Thereafter, the adsorbate was eluted with MnCl2-removed operation buffer 5 (FIG. 9). The resultant GnT-IV active fraction was pooled (Sample 6).
(vii) Superdex 200 Gel Chromatography
Sample 6 was concentrated with a small Q-sepharose FF column and applied to Superdex 200HR5/5 Column (1×30 cm; Pharmacia, Sweden) pre-equilibrated with operation buffer 6 (obtained by adding KCl to operation buffer 5 at a final concentration of 0.15 M) (FIG. 10). Operation buffer 6 was applied to the column at a flow rate of 0.25 ml/min to thereby obtain the GnT-IV active fraction (Sample 7).
(viii) The amount of protein, activity and specific activity in each purification step are summarized in Table 2. The final sample was purified 224,000-fold compared to the small intestine homogenate.
| TABLE 2 |
| |
| Purification of GnT-IV |
| |
Amount of |
Total enzyme |
|
|
Purification |
| |
protein |
activity |
Specific activity |
Yield |
factor |
| Purification Step |
(mg) |
(nmol/h) |
(nmol/h/mg) |
(%) |
(−fold) |
| |
| Bovine small intestine homogenate |
112,900 |
49,500 |
0.44 |
100 |
1 |
| Solubilized fraction |
24,100 |
14,500 |
0.60 |
29 |
1.4 |
| Q-Sepharose |
4,000 |
7,200 |
1.80 |
14 |
4.1 |
| Cu Chelate Sepharose |
450 |
3,670 |
8.10 |
7.4 |
18.4 |
| UDP-Hexanolamine I |
0.59 |
1,950 |
3,310 |
3.9 |
7,510 |
| UDP-Hexanolamine II |
0.035 |
1,420 |
40,600 |
2.9 |
92,200 |
| Superdex 200 |
0.008 |
790 |
98,800 |
1.6 |
224,000 |
| |
| Started from 2 kg of bovine small intestine. |
(3) Properties in Terms of Enzyme Chemistry and Protein Chemistry
(i) Purity
Sample 7 gave a single band at a molecular weight of 60 K in SDS-PAGE (FIG. 11). When Sample 7 was subjected to native-PAGE and the resultant band was cut out from the gel to determine GnT-IV activity, the location of the protein band agreed with the location of the activity (FIG. 12). Furthermore, any of GnT-I, -II -III or -V activity was not detected in Sample 7. From these findings, it was concluded that Sample 7 is pure GnT-IV. Taking into account that the apparent molecular weight of this protein was 77 K as determined by Triton X-100-containing gel filtration (FIG. 10), it is thought that GnT-IV does not have a subunit structure and expresses its activity in the form of a monomer. Whe n Sample 7 was treated wi th Peptide N-Glycosidase F (Boehringer-Mannheim, Germany), an increase in mobility was observed on SDS-PAGE. Thus, it is thought that GnT-IV from bovine small intestine is a glycoprotein having at least Asn-linked sugar chains.
(ii) Reaction Specificity
When this enzyme reacts on the GnT-II product type oligosaccharide represented by th e formula below as a substrate:
under the standard assay conditions, the enzyme yielded a single product (pyridylaminated oligosaccharide 1) as assayed by HPLC.
This product was collected, followed by determination of its structure by (i) a combination of Smith degradation and laser TOF-MS (time-of-flight mass spectrometer) and (ii)
1H-NMR. Thus, the reaction specificity of this enzyme was examined. When pyridylaminated oligosaccharide 1 was subjected to Smith degradation according to the method of Kobata and Takasaki [Kobata, A. and Takasaki, S. (1993) in Glycobiology “A Practical Approach” (Fukuda, M. and Kobata, A., eds) 165-185, IRL Press, Oxford, England], its mass number changed from 1599.0 to 795.30 as a result of the first degradation and further changed to 634.68 as a result of the second degradation. This agrees to the reaction pathway as shown in FIG.
13. Thus, it was concluded that the reaction product of this enzyme has the following structure:
Further, when
pyridylaminated oligosaccharide 1 was subjected to
1H-NMR, a peak of 4.53 ppm which corresponds to the anomeric proton of GlcNAc7 shown in the following formula was detected; its coupling constant J1, 2 was 7.9 Hz (FIG.
14). These results indicate that GlcNAc7 is, as shown in the formula below, attached to
position 4 of Man4 via β-type linkage, supporting the above structure completely.
(iii) Optimum pH
The optimum pH of this enzyme is around 7.5 as shown in FIG. 15.
(iv) Requirement of Divalent Cation
As shown in Table 3, this enzyme is deactivated by the addition of EDTA (ethylenediamine tetra-acetic acid). A divalent cation is essential for its activity. Among divalent cations, Mn2+ shows the greatest effect, followed by Co2+ and Mg2+. Weak effect is recognized in Ca2+ and Fe2+. The optimum concentration of Mn2+ is around 10 mM as shown in FIG. 16.
| TABLE 3 |
| |
| Divalent Cation Requirement of GnT-IV |
| |
None |
5.6 |
| |
EDTA |
0 |
| |
MnCl 2 |
100 |
| |
CoCl2 |
74.8 |
| |
MgCl2 |
72.5 |
| |
CaCl2 |
7.2 |
| |
FeCl2 |
9.8 |
| |
CuCl 2 |
0 |
| |
|
GnT-IV activity was determined by adding each of the metal ions (10 mM) to a GnT-IV sample from which metal ions had been removed. GnT-IV activity is represented in per cent in the Table, wherein the activity when 10 mM MnCl2 was added is regarded as 100%.
(v) Inhibition by Sugar Nucleotides
As shown in Table 4, UDP inhibited the activity of this enzyme most strongly. The inhibitory effects of UDP-glucose, UDP-GalNAc, 2′-deoxy-UDP and UDP-hexanolamine (Sigma, USA) followed that of UDP in this order. Uridine, UMP, TDP and CDP exhibited little inhibitory effect.
| TABLE 4 |
| |
| Inhibition of GnT-IV by Nucleotides |
| |
None |
100 |
| |
Uridine |
115 |
| |
UMP |
97.3 |
| |
UDP |
27.3 |
| |
UTP |
88.2 |
| |
TDP |
110 |
| |
CDP |
112 |
| |
2′-deoxy-UDP |
67.4 |
| |
UDP-hexanolamine |
73.6 |
| |
UDP-glucose |
56.6 |
| |
UDP-galactose |
87.3 |
| |
UDP-glucuronic acid |
92.3 |
| |
UDP-N-acetylgalactosamine |
59.7 |
| |
|
GnT-IV activity when each nucleotide (2 mM) was added in the presence of 0.5 mM UDP-GlcNAc is expressed in per cent in the Table, wherein the activity when nothing was added is regarded as 100%.
(vi) Substrate Specificity
As shown in Table 5, this enzyme preferred the GnT-V product type oligosaccharide (E in Table 5) the most as an acceptor. Next to this, the enzyme preferred the GnT-II product type oligosaccharides (D in Table 5).
When the reactivity of this enzyme toward the GnT-II type oligosaccharide is regarded as 100%, this enzyme exhibits reactivities of 0% and 54% toward the core type oligosaccharides (A in Table 5) and the GnT-I product type oligosaccharides (C in Table 5), respectively.
This enzyme exhibits a reactivity of 46% toward a structure of GnT-II product type oligosaccharide in which fucose is attached via α1→6 linkage to the GlcNAc at the reducing terminus (F in Table 5).
This enzyme exhibits a reactivity of 0% toward a structure of GnT-II product type oligosaccharides in which the GlcNAc on the α1→3 mannose is lacking (B in Table 5).
This enzyme exhibits a reactivity of 16% toward a structure of GnT-II product type oligosaccharides in which galactose is attached via β1→4 linkage to the GlcNAc on the α1→6 mannose (G in Table 5), and a reactivity of 0% toward a structure of GnT-II product type oligosaccharide in which galactose is attached via β1→4 linkage to the GlcNAc on the α1→3 mannose (H and I in Table 5).
This enzyme exhibits a reactivity of 0% toward a structure of GnT-II product type oligosaccharides in which GlcNAc is attached via β1→4 linkage to the β1→4 mannose (J in Table 5).
The substrate specificity of this enzyme as described above almost agree with the substrate specificity of GnT-IV predicted by Schachter et al. [Glesson, P. A. and Schachter, H. (1983) J. Biol. Chem., 258, 6162-6173]. Thus, it has become clear that this enzyme of the invention is the very GnT-IV that has long been a missing link in the biosynthesis of complex type sugar chains.
| |
TABLE 5 |
| |
|
| |
|
Relative |
| |
|
Activity of |
| |
|
GlcNAc |
| |
Receptor Oligosaccharide |
Transfer (%) |
| |
|
| |
| A |
|
0 |
| |
| B |
|
0 |
| |
| C |
|
54 |
| |
| D |
|
100 |
| |
| E |
|
164 |
| |
| F |
|
46 |
| |
| G |
|
16 |
| |
| H |
|
0 |
| |
| I |
|
0 |
| |
| J |
|
0 |
| |
(vii) Kinetic Parameters
Under the assay conditions as described in Reference Example 2, the Km and Vmax values of this enzyme toward the GnT-II product type oligosaccharides were 0.73 mM and 3.23 μM/min, respectively, and these values toward the GnT-V product type oligosaccharides were 0.13 mM and 1.75 μM/min, respectively. The Km value toward the UDP-GlcNAc was 0.22 mM.
Among the pyridylaminated oligosaccharides obtained, those represented by the following formulas were found to be novel oligosaccharides:
(viii) Action on Glycoproteins
In order to demonstrate that GnT-IV can act on not only oligosaccharide substrates but also oligosaccharide chains on glycoproteins, GnT-Iv reacts on asialo agalacto glycoproteins using UDP-[14 C] GlcNAc as a sugar donor. Then, the reaction products were analyzed by SDS-PAGE and fluorography (Panels A and B, FIG. 17). As shown in lanes 2 and 5 of Panel B in FIG. 17, transfer of [14 C] GlcNAc to asialo agalacto human transferrin and asialo agalacto, CHO cell-derived recombinant human erythropoietin.
The human transferrin having the GnT-IV product type sugar chain (of the following formula) obtained by this GnT-IV reaction is a novel substance which does not occur in nature.
About 1 mg of this enzyme of the invention finally purified was electrophoresed on 0.1% SDS-10% polyacrylamide gel according to the method of Laemmli [Laemmli, U. K. Nature (1970) 313, 756-762]. The separated proteins were electroblotted onto a PVDF membrane. The protein fixed on the membrane was S-carboxymethylated and then digested with lysylendopeptidase Achromobacter protease I (AP-I) (Wako Pure Chemical Industries, Ltd) to obtain an AP-I-digested fragment mixture. The AP-I-digested PVDF membrane was further digested with Asp-N (Takara Shuzo) to obtain an Asp-N-digested fragment mixture. Each of the peptide fragment mixtures was separated by high performance liquid chromatography and subjected to amino acid sequence analysis. As a result, the sequences shown in SEQ ID NOS: 1-14 were obtained.
EXAMPLE 3
Isolation and Identification of Bovine GnT-IVa cDNA
(1) RT-PCR
Based on the amino acid sequences shown in SEQ ID NOS: 7 and 11 obtained in Example 2, oligomer AP-5F shown in SEQ ID NO: 15 and oligomer DN-9R shown in SEQ ID NO: 16 were synthesized respectively. An RT-PCR was performed using as a template the total RNA extracted from bovine small intestine tissue by the guanidium isothiocyanate method and using the above primers. As a result, an amplified fragment of about 170 bp which seemed specific was obtained. This fragment was subcloned.
(2) Screening of a Library
A bovine small intestine cDNA library (Clontech, USA) was screened with the above-mentioned RT-PCR product to obtain four positive plaques. The nucleotide sequences of these clones were determined. The resultant sequences contained a number of nucleotide sequences coding for some of the partial amino acid sequences (SEQ ID NOS: 1-14) obtained in Example 2, and also contained a sequence appearing to be a termination codon. Using a fragment of 150 bp representing the most upstream region of the resultant nucleotide sequence, the library was screened again to obtain two positive plaques. The nucleotide sequences of these clones were determined. Then, the library was further screened similarly with a probe of 150 bp of the most upstream region, however, new clones were not obtained.
(3) 5′ RACE (Rapid Amplification of the cDNA Ends)
Subsequently, 5′ RACE was performed in order to obtain a full-length cDNA. Using the sequence of the most upstream region obtained by the phage screening, the first 5′ RACE was performed. However, an initiation codon could not be found. Then, based on the sequence obtained by the first 5′ RACE, the second 5′ RACE was performed to thereby obtain a sequence containing an initiation codon. This sequence was ligated to the previously obtained partial gene sequence of the phage clone to thereby obtain a gene fragment containing an intact open reading frame (Gene 1). The nucleotide sequence for the thus obtained gene fragment is shown in SEQ ID NO: 17, and the amino acid sequence deduced therefrom is shown in SEQ ID NO: 18. It was confirmed that this DNA fragment contains all of the nucleotide sequences coding for the 14 partial amino acid sequences (SEQ ID NOS: 1-14) obtained in Example 2.
EXAMPLE 4
Construction of an Expression Vector using the Cloned Bovine GnT-IVa Gene and a Method for Producing GnT-IVa Enzyme
(1) Construction of a Vector
A primer (SEQ ID NO: 19) which introduces an XhoI site into a region upstream of the initiation codon of Gene 1 and another primer (SEQ ID NO: 20) which introduces an XbaI site into a region downstream of the termination codon of the gene were synthesized. Then, the entire gene coding for GnT-IV enzyme was amplified by PCR with the primers. The amplified fragment obtained was digested with XhoI and XbaI, and inserted between the XhoI and XbaI sites of pSVL vector (Pharmacia, Sweden) to prepare plasmid pBGT4.
(2) Introduction into COS7 Cells
Plasmid pBGT4 was introduced into COS7 cells (RIKEN Cell Bank) by electroporation. Briefly, 10 μg of the plasmid was added to approximately 5×106 cells in 0.8 ml of PBS(−) (Nissui Pharmaceutical Co.). A voltage of 1600 V was applied at a capacitance of 25 μF with a gene pulser (BioRad, USA) at room temperature to introduce the gene into the cells. The resultant cells were transferred to a 90 mm laboratory dish and cultured in 10 ml of Dulbecco's modified Eagle's medium (Base Catalogue No. 12430, Life Technologies, Inc., USA) containing 10% fetal bovine serum under 5% CO2 at 37° C. for 72 hours. Thereafter, the cells were recovered and suspended in 100 μl of a buffer (5 mM Tris-HCl, pH 7.5, 2 mM MgCl2, 1 mM phenylmethylsulfonyl fluoride), followed by sonication and centrifugation at 2000×g for 5 minutes. Thus, a cell extract was obtained.
(3) Assay of GnT-IV Activity
GnT-IV activity in the cell extract was determined by the method described in Reference Example 2. The results are shown in Table 6. Compared to the extract from cells into which PSVL vector was introduced as a control, the extracts into which plasmid pBGT4 was introduced exhibited 44-78 times higher GnT-IV activity per cell. From these results, it was confirmed that Gene 1 encodes GnT-IV enzyme. Thus, GnT-IV enzyme can be produced by cultured cells according to this method.
| |
TABLE 6 |
| |
|
| |
|
Specific Activity |
|
| |
Plasmid |
(pmol/hr/mg protein) |
Activity Ratio |
| |
|
| |
| |
pSVL |
409 |
1 |
| |
pBGT4(#1) |
29623 |
72 |
| |
pBGT4(#2) |
31773 |
78 |
| |
pBGT4(#3) |
20182 |
44 |
| |
|
| |
Reaction time: 4 hours |
| |
Activity ratios are expressed in relation to the total activity of pSVL that is regarded as 1. |
EXAMPLE 5
Isolation and Identification of Human GnT-IVa cDNA
(1) RT-PCR
Based on the nucleotide sequence of bovine GnT-IVa obtained in Example 3, primer h1-2F shown in SEQ ID NO: 21 and primer h1-1R shown in SEQ ID NO: 22 were synthesized. Using total RNA from human liver (Clontech, USA) as a template, an RT-PCR was performed with the above primers. As a result, an amplified fragment of about 650 bp which seemed to be specific was obtained. This fragment was subcloned, and the nucleotide sequence thereof was determined.
(2) screening of a Library
A CDNA library from human liver (Clontech, USA) was screened using the 685 bp DNA fragment obtained by the above RT-PCR as a probe.
Two positive plaques of hGT4/λgt10-1 and hGT4/λgt10-2 were obtained. The nucleotide sequences of the inserts in these phage clones were determined. As a result, hGT4/λgt10-1 contained a 804 bp DNA region and hGT4/λgt10-2 contained a 2115 bp DNA region. The former region was entirely included in the latter region. As shown in SEQ ID NO: 23, the DNA fragment contained in hGT4/λgt10-2 had an open reading frame (ORF) highly homologous to the amino acid sequence of bovine GnT-IVa (96% identical). From the results described in Example 6, it was confirmed that this ORF is human GnT-IVa gene. The amino acid sequence of this ORF is shown in SEQ ID NO: 24.
EXAMPLE 6
Construction of an Expression Plasmid for Human GnT-IVa Gene and a Method for Producing Human GnT-IVa Enzyme
(1) Construction of Expression Plasmid pHGT4-1 for Human GnT-IVa Gene
A primer (h1-7F; SEQ ID NO: 25) which introduces an XhoI site into a region upstream of the initiation codon of human GnT-IVa gene and another primer (h1-7R; SEQ ID NO: 26) which is complementary to a region downstream of the termination codon of the gene were synthesized. Using RNA from human liver (Clontech, USA) as a template, the entire gene coding for human GnT-IVa enzyme was amplified by RT-PCR with the above primers. The resultant amplified fragment was inserted in the SrfI site of plasmid pCRScript Amp SK(+) (Stratagene, DNA) in the opposite direction to the transcription of lacZ gene. Using the resultant plasmid, it was confirmed by nucleotide sequence analysis that the amplified fragment encodes the amino acid sequence shown in SEQ ID NO: 24. Further, this plasmid was digested with XhoI and SacI to obtain an XhoI-SacI 1.7 kb fragment. This fragment was inserted bewteen the XhoI and Sacd sites of pSVL vector (Pharmacia, Sweden) to prepare an expression plasmid pHGT4-1 for human GnT-IVa gene.
(2) Introduction of Human GnT-IVa Gene into COS7 Cells
Plasmid pHGT4-1 was introduced into COS7 cells by electroporation. The resultant cells were cultured under 10% CO2 at 37° C. for 72 hours. Then, the cells were harvested, suspended in 100 μl of a buffer (5 mM Tris-HCl, pH 7.5, 2 mM magnesium chloride, 1 mM phenylmethylsulfon yl fluoride), disrupted by sonication, centrifuged at 2000×g for 5 minutes and collect supernatant to obtain a cell extract.
(3) Expression of Human GnT-IVa Gene in COS7 Cells
GnT-IV activity in the cell extract was determined by the method described in Reference Example 2. The results are shown in Table 7. Compared to the extract from cells into which pSVL vector was introduced as a control, the extracts from cells into which plasmid pHGT4-1 was introduced exhibited 21-28 times higher GnT-IV activity per cell. From these results, it was confirmed that GnT-IVa gene shown in SEQ ID NO: 23 encodes the glycosyltransferase GnT-IV. It was also confirmed that human GnT-IVa enzyme can be produced by cultured cells according to this method.
| |
TABLE 7 |
| |
|
| |
|
Specific Activity |
|
| |
Plasmid |
(pmol/hr/mg protein) |
Activity Ratio |
| |
|
| |
| |
pSVL |
1037 |
1 |
| |
pHGT4-1(#1) |
28951 |
28 |
| |
pHGT4-1(#2) |
21788 |
21 |
| |
pHGT4-2(#1) |
11024 |
11 |
| |
pHGT4-2(#2) |
8029 |
8 |
| |
|
| |
Reaction time: 1.3 hours |
| |
Activity ratios are expressed in relation to the total activity of pSVL that is regarded as 1. |
Reaction time: 1.3 hours
Activity ratios are expressed in relation to the total activity of pSVL that is regarded as 1.
EXAMPLE 7
Isolation and Identification of Human GnT-IVb cDNA
(1) Acquisition of Human GnT-IVa-like Gene by PCR, RT-PCR and 5′ RACE (Rapid Amplification of CDNA Ends)
Nucleotide sequences having similarity to the nucleotide sequence of human GnT-IVa gene obtained in Example 3 were searched for in the DNA database GenBank by BLASTN. As a result, Accession Numbers R12057, H10557 and W16571 were found out. Then, primer h2-45F shown in SEQ ID NO: 27 and primer h2-43R shown in SEQ ID NO: 28 were synthesized to perform a PCR using a cDNA library from human brain of Quick Screen Human cDNA Library Panel (Clontech, USA) as a template. The amplified fragment was subcloned into the SrfI site of pCRScript Amp SK(+) (Stratagene, USA) and subjected to analysis of the nucleotide sequence. Also, primer h2-2F shown in SEQ ID NO: 29 and primer h2-1R shown in SEQ ID NO: 30 were synthesized to perform an RT-PCR using total RNA from human lung (Clontech, USA) as a template. As a result, an amplified fragment of about 500 bp of the expected size was obtained. Then, this fragment was subcloned into the SrfI site of pCRScript Amp SK(+) (Stratagene, USA) and subjected to analysis of the nucleotide sequence.
The thus obtained nucleotide sequences of the two DNA fragments were overlapping with each other forming a region of 1006 bp. In this region, one reading frame which encodes the homologous amino acid sequences to those of bovine and human GnT-IVa was recognized. Thus, the existence of a protein relating to GnT-IVa proteins was suggested.
Then, possible nucleotide sequences which may be an upstream sequence to R12057 or a downstream sequence to W16571 were searched for in the DNA database GenBank by BLASTN. As a result, R15554 was found as an upstream sequence to R12057, and W16466 as a downstream sequence to W16571. However, a apparently inappropriate termination codon was contained in the ORFs deduced from these nucleotide sequences. Therefore, in order to confirm the nucleotide sequences, DNA fragments were obtained by RT-PCR. As primers, h2-1F shown in SEQ ID NO: 31, h2-3F shown in SEQ ID NO: 32 and h2-8R shown in SEQ ID NO: 33 were synthesized. With a combination of h2-1F and the h1-1R described in Example 5, or a combination of h2-3F and h2-8R, an RT-PCR was performed using total RNA from human liver (Clontech, USA) as a template. Amplified fragments of about 550 bp and about 300 bp, both coinciding with the expected sizes, were detected. Each of these fragments was subcloned in the SrfI site of pCRScript Amp SK(+) to analyze the nucleotide sequence thereof. As a result, it was confirmed that these fragments are respectively overlapping with an upstream region and a downstream region to the 1006 bp region between h2-45F and h2-1R mentioned above. In the ligated region of 1361 bp, an ORF was found which encodes 433 amino acids protein having high similarity to the amino acid sequences of bovine and human GnT-IVa proteins.
However, when this ORF is compared to the amino acid sequences of GnT-IVa proteins, it was presumed that the starting methionine should be present in a region upstream to this ORF. Therefore, the upstream region was obtained by 5′ -RACE using Human Lung 5′ -RACE-Ready cDNA (Clontech, USA). In the first PCR, an anchor primer and h2-5R shown in SEQ ID NO: 34 were used as primers. In the second PCR, an anchor primer and h2-3R shown in SEQ ID NO: 35 were used as primers. The fragments obtained by 5′ -RACE were purified, digested with ECORI and PstI, and then separated by agarose gel electrophoresis. A fragment of about 450 bp was recovered from the gel. This fragment was inserted between the EcoRI and PstI sites of pUC18 vector (Pharmacia, Sweden) to analyze the nucleotide sequence thereof. As a result, it was confirmed that this fragment is overlapping with a region upstream of the region between h2-1F and h2-8R. In the ligated region of 1758 bp, one ORF was confirmed which encodes 548 amino acids protein having high similarity to the amino acid sequences of bovine and human GnT-IVa proteins. The nucleotide sequence of this ORF is shown in SEQ ID NO: 36, and the amino acid sequence thereof in SEQ ID NO: 37. From the results described in Example 8 below, it was confirmed that this gene is human GnT-IVb gene.
EXAMPLE 8
Construction of an Expression Plasmid for Human GnT-IVb Gene and a Method for Producing Human GnT-IVb Enzyme
(1) Construction of Expression Plasmid pHGT4-2 for Human GnT-IVb Gene
A primer (h2-4: SEQ ID NO: 38) which introduces an XhoI site into a region upstream of the initiation codon of human GnT-IVb gene, and another primer (h2-10R: SEQ ID NO: 39) which introduces an XbaI site in a region downstream to the termination codon of the above gene were synthesized. Using these primers, the entire ORF coding for human GnT-IVb enzyme was amplified by RT-PCR with RNA from human lung (Clontech, USA) as a template. The amplified fragment was inserted into the SrfI site of plasmid pCRScript Amp SK(+), followed by determination of the nucleotide sequence thereof. As a result, it was confirmed that the amplified fragment is coding for the amino acid sequence of SEQ ID NO: 37. Further, this plasmid was digested with XhoI and XbaI to obtain an XhoI-XbaI 1.7 kb fragment. This fragment was inserted between the XhoI and XbaI sites of pSVL vector (Pharmacia, Sweden) to construct an expression plasmid pHGT4-2 for human GnT-IVb gene.
(2) Introduction of Human GnT-IVb Gene into COS7 Cells
Plasmid pHGT4-2 was introduced into COS7 cells by electroporation. The resultant cells were cultured under 10% CO2 at 37° C. for 72 hours. Then, the cells were recovered, suspended in 100 μl of a buffer (5 mM Tris-HCl, pH 7.5, 2 mM magnesium chloride, 1 mM phenyl-methylsulfonyl fluoride), disrupted by sonication, centrifuged at 2000×g for 5 minutes and collect supernatant to thereby obtain a cell extract.
(3) Expression of Human GnT-IVb Gene in COS7 Cells
GnT-IV activity in the cell extract was determined by the method described in Reference Example 2. The results are shown in Table 7 above. Compared to the extract from cells into which pSVL vector was introduced as a control, the extracts from cells into which plasmid pHGT4-2 was introduced exhibited 8-11 times higher GnT-IV activity per cell. From these results, it was confirmed that the GnT-IVb gene shown in SEQ ID NO: 36 encodes the glycosyltransferase GnT-IV. It was also confirmed that human GnT-IVb enzyme can be produced by cultured cells according to this method.
EXAMPLE 9
Construction of Expression Plasmids for Bovine GnT-IVa N-Terminal Deletion Mutants and their Expression of
(1) Construction of Expression Plasmids pSigIle93, pSigPro113 and pSigPro142 for Bovine GnT-IVa
A primer (XhoEsig: SEQ ID NO:40) which introduces an XhoI site into a region upstream of the signal sequence of human erythropoietin (GenBank Accession Number X02157) and an antisense primer (E4-1R: SEQ ID NO:41) which ligates the C-terminus of the above signal sequence to a part of the bovine GnT-IVa amino acid sequence spanning from position 93 (Ile) to the end were synthesized to amplify the signal sequence of human erythropoietin by PCR. Also, a sense primer (E4-1F: SEQ ID NO:42) corresponding to the above antisense primer and a primer (4EXPR: SEQ ID NO:20) which introduces an XbaI site in a region downstream of the termination codon of bovine GuT-IVa gene were synthesized to amplify a partial sequence of bovine GnT-IVa gene by PCR. Using portions of the resultant two PCR products as a mixed template, a PCR was performed with primers XhoEsig and 4EXPR. The amplified fragment was digested with XhoI and XbaI and inserted between the XhoI and XbaI sites of pSVL vector (Pharmacia, Sweden), to thereby construct plasmid pSigIle93 that expresses an amino acid sequence in which the human erythropoietin signal is linked to a part of the bovine GnT-IVa amino acid sequence spanning from position 93 to the end.
Plasmid pSigPro113 that expresses an amino acid sequence in which the human erythropoietin signal is linked to a part of the bovine GnT-IVa amino acid sequence spanning from position 113 (Pro) to the end; or plasmid pSigProl42 that expresses an amino acid sequence in which the human erythropoietin signal is linked to a part of the bovine GnT-IVa amino acid sequence spanning from position 142 (Pro) to the end was constructed respectively in the same manner as described above using E4-2R primer (SEQ ID NO:43) or E4-3R primer (SEQ ID NO:44) instead of E4-1R primer; and E4-2F primer (SEQ ID NO:45) or E4-3F primer (SEQ ID NO:46) instead of E4-1F primer.
(2) Introduction of plasmids expressing Bovine GnT-IVa N-Terminal Deletion Mutants into COS7 Cells
Plasmid pSigIle93, pSigPro113 or pSigPro142 was introduced into COS7 cells by electroporation. The resultant cells were cultured under 10% CO2 at 37° C. for 72 hours. Then, the cells and the culture supernatant were recovered separately. The cells were suspended in 100 μl of a buffer (5 mM Tris-HCl, pH 7.5, 2 mM magnesium chloride, 1 mM phenylmethylsulfonyl fluoride), disrupted by sonication and centrifuged at 2000×g for 5 minutes to thereby obtain a cell extract. The culture supernatant was concentrated to about 100 μl with Centriplus 30 (Amicon).
(3) Expression of Bovine GnT-IVa N-Terminal Deletion Mutants in COS7 Cells
GnT-IV activity in the culture supernatant and the cell extract was determined by the method described in Reference Example 2. The results are shown in Table 8. Compared to the total activity (i.e., activity in cells+activity in supernatant) of the cells into which pBGT4 vector was introduced as a positive control, the total activity of the cells into which pSigIle93 was introduced was more than 30%. Furthermore, more than one third of the activity was secreted into the culture supernatant. From these results, it was found that the amino acids from the N-terminus to position 92 of the bovine GnT-IVa amino acid sequence can be deleted while retaining the enzyme activity. It was also shown that GnT-IVa enzyme can be expressed secretively by using an appropriate secretion signal.
| TABLE 8 |
| |
| |
|
|
Activity |
Total |
| |
|
|
Ratio in |
Activity |
| |
|
Activity |
Each Fraction |
Ratio |
| Plasmid |
Fraction |
(pmol/hr) |
(%) |
(%) |
| |
| |
| pSVL |
Supernatant |
136 |
0.5 |
1.9 |
| pSVL |
Cells |
384 |
1.4 |
| pBGT4 |
Supernatant |
722 |
2.7 |
100.0 |
| pBGT4 |
Cells |
26152 |
97.3 |
| pSigIle93 |
Supernatant |
3106 |
11.6 |
31.9 |
| pSigIle93 |
Cells |
5471 |
20.4 |
| pSigPro113 |
Supernatant |
312 |
1.2 |
3.4 |
| pSigPro113 |
Cells |
606 |
2.3 |
| pSigPro142 |
Supernatant |
219 |
0.8 |
2.2 |
| pSigPro142 |
Cells |
381 |
1.4 |
| |
| Reaction time: 2.5 hours |
| The activity ratios are expressed in percent in relation to the total activity of pBGT4 that is regarded as 100%. |
EXAMPLE 10
Construction of Expression Plasmids for Bovine GnT-IVa C-Terminal Deletion Mutants and their Expression
(1) Construction of Expression Plasmids pCGly499, pCPro465, pCLys432 and pCPro383 for Bovine GnT-IVa
A primer (SEQ ID NO:19) which introduces an XhoI site into a region upstream of the initiation codon of bovine GnT-IVa gene and a primer (CGly499: SEQ ID NO:47) which ligates the termination codon after the Gly codon at position 499 and introduce an XbaI site in a region downstream to the termination codon above were synthesized to amplify a partial sequence of bovine GnT-IVa gene by PCR. The amplified fragment was digested with XhoI and XbaI, and inserted between the XhoI and XbaI sites of pSVL vector (Pharmacia, Sweden). Thus, plasmid pCGly499 which expresses the bovine GnT-IVa amino acid sequence up to position 499 (Glycine) was constructed. Using CPro465 primer (SEQ ID NO:48), CLys432 primer (SEQ ID NO:49) or CPro383 primer (SEQ ID NO:50) instead of CGly499 primer, three additional plasmids were constructed in the same manner. They were designated pCPro465 (plasmid which expresses the bovine GnT-IVa amino acid sequence up to position 465 (Proline)); pCLys432 (plasmid which expresses the bovine GnT-IVa amino acid sequence up to position 432 (Lysine)); and pCPro383 (plasmid which expresses the bovine GnT-IVa amino acid sequence up to position 383 (Proline)), respectively.
(2) Introduction of plasmids expressing Bovine GnT-IVa C-Terminal Deletion Mutants into COS7 Cells
Plasmid pCGly499, pCPro465, pCLys432 or pCPro383 was introduced into COS7 cells by electroporation. The resultant cells were cultured under 10% CO2 at 37° C. for 72 hours. Then, the cells were recovered and suspended in 100 μl of a buffer (5 mM Tris-HCl, pH 7.5, 2 mM magnesium chloride, 1 mM phenylmethylsulfonyl fluoride), disrupted by sonication and centrifuged at 2000×g for 5 minutes to thereby obtain a cell extract.
(3) Expression of Bovine GnT-IVa C-Terminal Deletion Mutants in COS7 Cells
GnT-IV activity in the cell extract was determined by the method described in Reference Example 2. The results are shown in Table 9. Compared to the GnT-IV activity of the extract from cells into which PBGT4 vector was introduced as a positive control, that activity of the extract from cells into which pCGly499, pCPro465, pCLys432 or pCPro383 was introduced was 15.2%, 12.1%, 2.8% or 104.2% per cell, respectively. From these results, it was shown that GnT-IV activity can be retained even if the amino acids from position 384 to the C-terminus in the bovine GnT-IVa amino acid sequence are deleted.
| |
TABLE 9 |
| |
|
| |
|
Specific Activity |
Activity Ratio |
| |
Plasmid |
(pmol/hr/mg protein) |
(%) |
| |
|
| |
| |
pSVL |
77 |
1 |
| |
pBGT4 |
14917 |
100 |
| |
pCGly499 |
2263 |
15 |
| |
pCPro465 |
1798 |
12 |
| |
pCLys432 |
410 |
3 |
| |
pCPro383 |
15551 |
104 |
| |
|
| |
Reaction Time: 2 hours |
| |
Activity ratios are expressed in percent in relation to the total activity of pBGT4 which is regarded as 100%. |
EXAMPLE 11
Construction of Plasmids to Express various GnT-IV Genes in E. coli and their Expression
(1) Construction of E. coli Expression Plasmid for Bovine GnT-IVa
A primer (BSP-N: SEQ ID NO:51) which introduces a BspHI site in a region upstream of the initiation codon of bovine GnT-IVa gene and another primer (C-Hd: SEQ ID NO:52) which introduces a HindIII site in a region downstream of the termination codon were synthesized to amplify the entire open reading frame of bovine GnT-IVa gene by PCR. The amplified fragment was digested with BspHI and HindIII, and introduced between the NcoI and HindIII sites of pTrc99A vector (Pharmacia, Sweden) to thereby construct plasmid pEBGT4. Using BSP-sN primer (SEQ ID NO:53) instead of BSP-N primer together with C-Hd primer, plasmid pEIle93 was constructed in a similar manner. Further, a gene coding for the open reading frame in which His-Tag is added to the C-terminus was amplified using BSP-N primer, a primer (CH-Hd: SEQ ID NO:54) which can introduce His-Tag, a termination codon and a HindIII site in a region downstream of the C-terminus of bovine GnT-IVa gene and a primer (H-Hd: SEQ ID NO:55) which has His-Tag, a termination codon and a HintdIII site, to thereby construct plasmid pEBGT4+His in a similar manner.
(2) Construction of E. coli Expression Plasmid for Human GnT-IVa Gene and Human GnT-IVb Gene
A primer (4aBSPIL94: SEQ ID NO:56) which introduces an initiation codon and an Ile codon in a region upstream of position 94 (Leu) of the human GnT-IVa amino acid sequence and which can also introduce a BSPHI site at a region further upstream thereof; a primer (4aCH-Hd: SEQ ID NO: 57) which introduces His-Tag, a termination codon and a HindIII site in a region downstream of the C-terminal amino acid; and H-Hd primer were synthesized to amplify a gene fragment composed a partial sequence of the human GnT-IVa amino acid sequence to which a sequence encoding His-Tag is added. The amplified fragment was digested with BspHI and HindIII, and inserted between the Nco I and HindIII sites of pTrc99A vector (Pharmacia, Sweden) to thereby construct plasmid pMA4a+His. Further, using CP383H-Hd primer (SEQ ID NO:58) instead of 4aCH-Hd primer, plasmid pCore+His was constructed in a similar manner (FIG. 18). A fragment of human GnT-IVb gene was amplified using a primer (4bBSP-N: SEQ ID NO:59) which introduces a Bsp HI site in a region upstream of the initiation codon of human GnT-IVb gene and 4bSACR primer (SEQ ID No: 60), digested with BspHI and Sac I, and then inserted between the Nco I and Sac I sites of pTrc99A vector (Pharmacia, Sweden). Between the SacI and HindIII sites of the resultant plasmid, a partial length of human GnT-IVb gene amplified using 4bSACF primer (SEQ ID NO:61), a primer (4bCH-Hd: SEQ ID NO: 62) which introduces His-Tag at the C-terminus of the human GnT-IVb amino acid sequence and H-Hd primer, and digested with Sac I and HindIII was inserted to thereby achieve plasmid pEHGT4-2+His. Further, a partial sequence of human GnT-IVb gene was amplified using a primer (4bNCOG91: SEQ ID NO:63) which introduces an Nco I site and an initiation codon in a region upstream of position 91 (Gly) of the human GnT-IVb amino acid sequence, 4bCH-Hd primer and H-Hd primer, digested with Nco I and HindIII, and then inserted between the Nco I and HindIII sites of pTrc99A vector (Pharmacia, Sweden) to thereby construct plasmid pMA4b+His.
(3) Introduction of Each Expression Plasmid into E. coli BL21 Strain
Each expression plasmid was introduced into competent cells of E. coli BL21 strain prepared by the calcium method. The resultant cells were cultured on LB agar plate containing 100 μg/ml ampicillin. The resultant colonies of E. coli transformed with each plasmid were inoculated into LB liquid medium and cultured under shaking at 37° C. overnight. Then, the culture was inoculated into a fresh LB liquid medium to give a concentration of 2%. While the turbidity (OD 595 nm) of the culture fluid was about 0.1 to 0.2, IPTG (isopropyl b-D-thiogalactopyranoside) was added thereto to give a final concentration of 1 mM. The cells were cultured at 37° C. for 2 hours or at 25° C. for 4 hours. Then, 500 μl of the cells was harvested. The cell pellet was suspended in 50 μl of a buffer (5 mM Tris-HCl, pH 7.5, 2 mM MgCl2, 1 mM phenylmethylsulfonyl fluoride), disrupted by sonication and centrifuged at 2000×g for 5 minutes to obtain a cell extract as a supernatant.
(4) Expression of Each Expression Plasmid in E. coli BL21 Strain GnT-IV activity in the cell extract was determined by the method described in Reference Example 2. Table 10 shows the results of the expression of the bovine gene. Although the extract from E. coli cells into which pTrc99A vector was introduced as a control had little GnT-IV, the extract from E. coli cells into which pEBGT4 was introduced had definite GnT-IV activity. From these results, it was demonstrated that GnT-IV enzyme can be produced by E. coli. The His-tag sequence added to the C-terminus of bovine GnT-IVa did not influence greatly upon GnT-IV activity. Thus, it was shown that an appropriate tag sequence can be added to GnT-IV enzyme. The mutant (pEIle93) in which the N-terminal 92 amino acid are deleted exhibited stronger GnT-IV activity. Thus, the expression of variants of GnT-IV enzyme which was confirmed in animal cells was also shown possible in E. coli.
| |
TABLE 10 |
| |
|
| |
|
Activity |
Activity Ratio |
| |
Plasmid |
(pmol/hr/mg protein) |
(%) |
| |
|
| |
| |
pTrc99A |
0 |
0 |
| |
pEBGT4 |
4611 |
100 |
| |
pEBGT4 + His |
3090 |
67 |
| |
pEIle93 |
5841 |
127 |
| |
|
| |
Reaction time: 3.0 hours |
| |
After IPTG addition, cells were cultured at 37° C. for 2 hours. |
| |
Activity ratios are expressed in percent in relation to the total activity of pEBGT4 which is regarded as 100%. |
Table 11 shows the results of the expression of the human gene. Compared to the extract from E. coli cells into which pTrc99A vector was introduced as a control, the extract from E. coli cells into which any of the expression plasmids was introduced had GnT-IV activity significantly. As shown in bovine GnT-IVa enzyme, it was also possible in human GnT-IVa and GnT-IVb enzymes to delete an N-terminal sequence while retaining the activity. Further, the human GnT-IVa enzyme which has both N-terminal deletion and C-terminal deletion exhibited high GnT-IV activity (pCore+His). This shows that those portions deleted in this mutant are not essential for GnT-IV activity.
| |
TABLE 11 |
| |
|
| |
|
Activity |
Activity Ratio |
| |
Plasmid |
(pmol/hr/mg protein) |
(%) |
| |
|
| |
| |
pTrc99A |
0 |
0 |
| |
pEBGT4 + His |
21390 |
637 |
| |
pMA4a + His |
3359 |
100 |
| |
pCore + His |
39766 |
1184 |
| |
pEHGT4-2 + His |
270 |
8 |
| |
pMA4b + His |
2812 |
84 |
| |
|
| |
Reaction time: 4.0 hours |
| |
After IPTG addition, cells were cultured at 25° C. for 4 hours. |
| |
Activity ratios are expressed in percent in relation to the total activity of pMA4a + His which is regarded as 100%. |
EXAMPLE 12
Conversion of the Sugar Chain Branching Structure of a Recombinant Erythropoietin (EPO) by Introducing Bovine or Human, GnT-IVa Gene into EPO-Producing CHO Cells
(1) Introduction of GnT-IVa Expression Plasmid into EPO-Producing CHO Cells
EPO-producing CHO cell clones were created according to the method disclosed in Japanese Examined Patent Publication.No. 2-17156. GnT-IVa expression plasmid pBGT4 or pHGT4-1 was introduced into the resultant cell clones Mo1 and H-5 by electroporation. In the introduction, 15 μg of the expression plasmid and 1.5 μg of a drug resistance marker plasmid (pSv2bsr from Kaken Pharmaceutical or pMAMneo from Clontech) were used in mixture. The electroporated cells were cultured under 10% CO2 at 37° C. for about 60 hours. Then, blasticidin S (Kaken Pharmaceuticals) (final concentration: 10 μg/ml) or geneticin (Life Technologies, Inc.) (final concentration: 500 μg/ml) was added to the medium, in which the cells were cultured for another 10 days to 2 weeks. Thus, clones resistant to either of two drugs were isolated.
(2) Confirmation of the Expression of the Introduced GnT-IVa Genes in EPO-Producing CHO Cell Clones
EPO-producing CHO cell clones (initial clones) and individual drug resistant clones were cultured in an appropriate scale. Total RNA from each clone was purified. Then, RNA dot blot analysis was performed using a part of GnT-IVa gene as a probe to thereby examine the amount of GnT-IVa mRNA. Further, GnT-IV activity expressed in the initial clones and the drug resistant clones was determined by the assay described in Reference Example 2. Those clones which gave a strong signal in RNA dot blot analysis and yet exhibited higher GnT-IV activity than the initial clones were selected and used for EPO production. The selected clones had increased GnT-IV activity; for example, MO1 (bovine GnT-IVa) #36 exhibited about 104-fold increase over MO1 clone, and H-5(human GnT-IVa) #23 exhibited about 125-fold increase over H-5 clone.
(3) Production of EPO using GnT-IVa Gene-Introduced EPO-Producing CHO Cell Clones
EPO is expressed secretively into culture fluid. Then, EPO-producing CHO cell clones MO1 and H-5, and the above-mentioned clones MO1 (bovine GnT-IVa) #36 and H-5(human GnT-IVa) #23 were cultured in roller bottles. First, each clone was adhesion-cultured in a growth medium, and then 1.5×107 cells were transferred to a 850 cm2 roller bottle containing 200 ml of a growth medium. The cells were cultured under 10% CO2 at 37° C. for 3 days so that they adhered to the bottle uniformly. Thereafter, the growth medium was removed, and the cells were washed with PBS buffer. Then, 200 ml of a serum-free medium was added to the bottle, in which the cells were cultured under 10% CO2 at 37° C. for 7 days. Thereafter, the culture supernatant was recovered.
As a growth medium, D-MEM/F12 mixed medium supplemented with 5% fetal bovine serum, 290 mg/liter L-glutamic acid, 1×MEM non-essential amino acid solution and 100 nM methotrexate was used. As a serum-free medium, the above medium without fetal bovine serum was used. EPO contained in each of the serum-free culture supernatants was quantitatively determined by ELISA usig anti-human EPO antibody.
(4) Analysis of EPOs Produced by GnT-IVa Gene-Introduced or -Non-Introduced Clones Based on Their Sugar Chain Structures
A recombinant EPO does not exist as a single molecule on isoelectric focusing gel; it is mixture of molecules with various electric charges. Since the protein moiety does not vary, it has been shown that the difference in electric charge among these molecules is based on the difference in sugar chain structure; such mixture of molecules is called glycoforms [Watson, E. and Yao, F., Anal. Biochem. (1993), 210, 389-93]. EPO has three Asn-linked sugar chains; the branching structures of individual sugar chains vary from biantennary to tetraantennary. Gal (galactose) is further attaching to the end of each of branched GlcNAc's, and sialic acid is further attaching to this Gal. Therefore, if the degree of sugar chain branching is increased by the introduction of GnT-IVa gene, the number of sialic acid molecules attaching to Gal should increase and, thus, the content of glycoforms with low isoelectric point should increase. Then, the inventors performed analysis by isoelectric focusing to detect changes in the sugar chain structure of the EPOs produced by GnT-IVa gene-introduced EPO-producing cells.
For the isoelectric focusing, Multiphor II equipment manufactured by Pharmacia was used. The gel was composed of 5% acrylamide (30: 0.8) and 1.5% Pharmalyte 2.5-5 (Pharmacia). As the (+) electrode solution, 0.1 M sulfuric acid was used. As the (−) electrode solution, 0.2 M L-histidine was used. After the isoelectric focusing, samples were electrophoretically transferred onto a PVDF membrane, followed by Western blot analysis using anti-EPO mouse monoclonal antibody to detect the bands of individual glycoforms of the EPOs. Briefly, the serum-free culture supernatant of each cell clone was concentrated to about 7 to 1000-fold with Centriplus 30 and Microcon 30 (both manufactured by Amicon), if necessary. At the beginning, about 50-100 IU of EPO was used as a sample, but this amount was adjusted appropriately so that the intensities of bands detected by Western blot analysis would be almost equal between samples.
When the EPO from MO1 clone was compared to the EPO from MO1 (bovine GnT-IVa) #36 clone, it was confirmed that the positions of major glycoforms in the latter show a shift to the low pI side (+electrode solution side) by at least one glycoporm (FIG. 19). From this result, it is thought that the GnT-IVa enzyme expressed as a result of the gene introduction increased the number of GlcNAc branches in the Asn-linked sugar chains attaching to EPO, thus increasing the number of sialic acid molecules attaching to increase the content of glycoforms with low isoelectric points. A similar analysis was performed on H-5 clone and H-5(human GnT-IVa )#23 clone. As a result, it was also found that the positions of major EPO glycoforms in the latter shift to the low pI side (FIG. 19).
From the above, it was demonstrated that it is possible to modify the structure of Ans-linked sugar chains of the protein produced by the cell by introducing a GnT-IVa gene into any cell.
Industrial Applicability
According to the present invention, a novel β1→4 N-acetyl-glucosaminyltransferase (GnT-IV), a method for producing the GnT-IV enzyme and a gene coding for the GnT-IV are provided. With the GnT-IV of the present invention, it has become possible to produce a glycocoujugate having a branching structure which could not be formed with conventional glycosyltransferases. Thus, the GnT-IV of the invention is useful not only for producing or improving glycoconjugate type pharmaceuticals, reagents and foods, but also for modifying the sugar chain structure of any biopolymer.
The GnT-IV gene of the invention is also useful for diagnosing or treating diseases such as cancer and for modifying the sugar chain structure of glycoconjugate products produced by microorganisms.
Further, an antibody or anti-serum raised against the GnT-IV protein of the invention as an antigen, or a part or all of the GnT-IV gene of the invention as a probe is useful for characterizing microorganisms, cultured cells, various animal tissues, blood cells and blood or for diagnosing diseased cells or tissues such as cancer.
63
1
8
PRT
Bovine
1
Asp Asn Leu Tyr Pro Glu Glu Lys
1 5
2
11
PRT
Bovine
2
Asp Tyr Val Asn Gly Val Val Ala Asn Glu Lys
1 5 10
3
21
PRT
Bovine
3
Glu Ile Ser Ser Gly Leu Val Glu Ile Ile Ser Pro Pro Glu Ser Tyr
1 5 10 15
Tyr Pro Asp Leu Thr
20
4
8
PRT
Bovine
4
Glu Arg Val Arg Trp Arg Thr Lys
1 5
5
15
PRT
Bovine
5
Lys Gln Asn Leu Asp Tyr Cys Phe Leu Met Met Tyr Ala Gln Glu
1 5 10 15
6
6
PRT
Bovine
6
Asp His Ile Leu Trp Val
1 5
7
14
PRT
Bovine
7
Lys Ile His Val Asn Pro Pro Ala Glu Val Ser Thr Ser Leu
1 5 10
8
10
PRT
Bovine
8
Lys Val Tyr Gln Gly His Thr Leu Glu Lys
1 5 10
9
10
PRT
Bovine
9
Asp Phe Phe Trp Ala Ile Thr Pro Val Ala
1 5 10
10
6
PRT
Bovine
10
Asp Tyr Ile Leu Phe Lys
1 5
11
15
PRT
Bovine
11
Asp Lys Pro Val Asn Val Glu Ser Tyr Leu Phe His Ser Gly Asn
1 5 10 15
12
10
PRT
Bovine
12
Asp Ile Leu Leu Asn Thr Thr Val Glu Val
1 5 10
13
9
PRT
Bovine
13
Lys Ser Glu Gly Leu Asp Ile Ser Lys
1 5
14
8
PRT
Bovine
14
Asp Gly Tyr Phe Arg Ile Gly Lys
1 5
15
29
DNA
Artificial Sequence
Description of Artificial Sequence Primer
15
aaratycayg tbaaycchcc hgcngargt 29
16
35
DNA
Artificial Sequence
Description of Artificial Sequence Primer
16
tgraavarrt arswytcvac rttvacdggy ttrtc 35
17
2246
DNA
Bovine
CDS
(288)..(1892)
17
ggcggctgct cggtggcggc tcgtcggcgg ccgcggcagg actggcagcg ccggcggcgg 60
ggagaaagaa gcatccacct atgaagaccg tgcagacagt cctgaataat aattgtgaat 120
ggtgtggctg ccagactagt tctgctgagc atctgaaatg aacctctcct attgattgtt 180
tcagttggcc ccgagccagg agtactgggt ttgcttgact tcaggataaa aagaaacgga 240
cttggttatc atcgtaaaca tatgaaccag tgtgatggtg aaatgag atg agg ctc 296
Met Arg Leu
1
cga aat gga act gta gcc act gtt tta gca ttt atc acc tcg ttc ctc 344
Arg Asn Gly Thr Val Ala Thr Val Leu Ala Phe Ile Thr Ser Phe Leu
5 10 15
act tta tct tgg tat aca aca tgg caa aat ggg aaa gaa aaa gtg att 392
Thr Leu Ser Trp Tyr Thr Thr Trp Gln Asn Gly Lys Glu Lys Val Ile
20 25 30 35
gct tat caa cga gaa ttt ctt gct ctg aaa gaa cgt ctc cga ata gct 440
Ala Tyr Gln Arg Glu Phe Leu Ala Leu Lys Glu Arg Leu Arg Ile Ala
40 45 50
gaa cat cga atc tct cag cgc tct tct gag ctc agt gcc att gta cag 488
Glu His Arg Ile Ser Gln Arg Ser Ser Glu Leu Ser Ala Ile Val Gln
55 60 65
caa ttc aag cgt gta gaa gca gaa aca aac agg agt aag gat cca gtg 536
Gln Phe Lys Arg Val Glu Ala Glu Thr Asn Arg Ser Lys Asp Pro Val
70 75 80
aat aaa ttt tca gat gat acc cta aag ata cta aag gag tta aca agc 584
Asn Lys Phe Ser Asp Asp Thr Leu Lys Ile Leu Lys Glu Leu Thr Ser
85 90 95
aaa aag tct ctt caa gtg cca agt att tat tat cat ttg cct cat tta 632
Lys Lys Ser Leu Gln Val Pro Ser Ile Tyr Tyr His Leu Pro His Leu
100 105 110 115
ttg caa aat gaa gga agc ctt caa cct gcc gtg cag atc gga aat gga 680
Leu Gln Asn Glu Gly Ser Leu Gln Pro Ala Val Gln Ile Gly Asn Gly
120 125 130
cga aca gga gtt tca ata gta atg gga att cct aca gtg aag aga gaa 728
Arg Thr Gly Val Ser Ile Val Met Gly Ile Pro Thr Val Lys Arg Glu
135 140 145
gtt aaa tct tac ctc ata gaa act ctt cat tcc ctt att gat aat ctg 776
Val Lys Ser Tyr Leu Ile Glu Thr Leu His Ser Leu Ile Asp Asn Leu
150 155 160
tat cct gaa gag aag ttg gac tgt gtt ata gta gtc ttc ata gga gag 824
Tyr Pro Glu Glu Lys Leu Asp Cys Val Ile Val Val Phe Ile Gly Glu
165 170 175
aca gat act gat tat gta aat ggt gtt gta gcc aac ctg gag aaa gaa 872
Thr Asp Thr Asp Tyr Val Asn Gly Val Val Ala Asn Leu Glu Lys Glu
180 185 190 195
ttt tct aaa gaa atc agt tct ggc ttg gtg gaa ata ata tca cct cct 920
Phe Ser Lys Glu Ile Ser Ser Gly Leu Val Glu Ile Ile Ser Pro Pro
200 205 210
gaa agc tat tat cct gac ctg acg aac tta aag gag aca ttt gga gat 968
Glu Ser Tyr Tyr Pro Asp Leu Thr Asn Leu Lys Glu Thr Phe Gly Asp
215 220 225
tct aaa gaa aga gta aga tgg aga aca aag caa aac cta gat tat tgt 1016
Ser Lys Glu Arg Val Arg Trp Arg Thr Lys Gln Asn Leu Asp Tyr Cys
230 235 240
ttt cta atg atg tat gct cag gaa aaa ggc aca tac tac atc cag ctt 1064
Phe Leu Met Met Tyr Ala Gln Glu Lys Gly Thr Tyr Tyr Ile Gln Leu
245 250 255
gaa gat gat att att gtc aaa cag aat tac ttt aac acc ata aag aat 1112
Glu Asp Asp Ile Ile Val Lys Gln Asn Tyr Phe Asn Thr Ile Lys Asn
260 265 270 275
ttt gca ctt caa ctt tct tct gag gaa tgg atg ata ctt gag ttc tcc 1160
Phe Ala Leu Gln Leu Ser Ser Glu Glu Trp Met Ile Leu Glu Phe Ser
280 285 290
cag ctg gga ttc att ggt aaa atg ttt caa gca cct gac cca ctc ctg 1208
Gln Leu Gly Phe Ile Gly Lys Met Phe Gln Ala Pro Asp Pro Leu Leu
295 300 305
att gtg gaa ttc ata ttt atg ttc tat aag gag aag ccc atc gac tgg 1256
Ile Val Glu Phe Ile Phe Met Phe Tyr Lys Glu Lys Pro Ile Asp Trp
310 315 320
ctc ttg gac cat att ctg tgg gtc aaa gtc tgc aac ccg gaa aaa gat 1304
Leu Leu Asp His Ile Leu Trp Val Lys Val Cys Asn Pro Glu Lys Asp
325 330 335
gca aaa cac tgt gat cga cag aag gca aat ctg cga att cgt ttc aga 1352
Ala Lys His Cys Asp Arg Gln Lys Ala Asn Leu Arg Ile Arg Phe Arg
340 345 350 355
ccg tcc ctt ttc caa cac gtt ggt ctg cat tct tca ctc aca gga aaa 1400
Pro Ser Leu Phe Gln His Val Gly Leu His Ser Ser Leu Thr Gly Lys
360 365 370
att cag aaa ctc acg gat aaa gat tac atg aaa cca tta ctg ctc aaa 1448
Ile Gln Lys Leu Thr Asp Lys Asp Tyr Met Lys Pro Leu Leu Leu Lys
375 380 385
atc cat gta aac ccc cct gca gag gta tct act tct ttg aag gtc tac 1496
Ile His Val Asn Pro Pro Ala Glu Val Ser Thr Ser Leu Lys Val Tyr
390 395 400
caa ggt cat aca ctg gag aaa act tac atg ggt gag gac ttc ttc tgg 1544
Gln Gly His Thr Leu Glu Lys Thr Tyr Met Gly Glu Asp Phe Phe Trp
405 410 415
gct ata acc cca gta gct gga gac tac atc cta ttt aaa ttc gac aag 1592
Ala Ile Thr Pro Val Ala Gly Asp Tyr Ile Leu Phe Lys Phe Asp Lys
420 425 430 435
cca gtc aat gtg gaa agt tat ttg ttc cat agt ggc aac cag gat cat 1640
Pro Val Asn Val Glu Ser Tyr Leu Phe His Ser Gly Asn Gln Asp His
440 445 450
cca ggg gat att ctg ctc aac aca acg gtg gaa gtt ctg cct ttg aag 1688
Pro Gly Asp Ile Leu Leu Asn Thr Thr Val Glu Val Leu Pro Leu Lys
455 460 465
agt gaa ggt ttg gac atc agc aaa gaa acc aaa gac aaa cga tta gaa 1736
Ser Glu Gly Leu Asp Ile Ser Lys Glu Thr Lys Asp Lys Arg Leu Glu
470 475 480
gat ggc tat ttc aga ata ggg aaa ttt gaa aac ggt gtt gcg gaa ggg 1784
Asp Gly Tyr Phe Arg Ile Gly Lys Phe Glu Asn Gly Val Ala Glu Gly
485 490 495
atg gtg gat ccc agc cta aac ccc att tcg gcc ttc cga ctt tca gtt 1832
Met Val Asp Pro Ser Leu Asn Pro Ile Ser Ala Phe Arg Leu Ser Val
500 505 510 515
att cag aat tct gct gtt tgg gcc att ctt aat gag atc cat att aaa 1880
Ile Gln Asn Ser Ala Val Trp Ala Ile Leu Asn Glu Ile His Ile Lys
520 525 530
aaa gtc aca aac tgaccatctc tactaagaaa ccaacacatt ttttccctgt 1932
Lys Val Thr Asn
535
gaatttgttg attaaagaca gctgagcacg tacctttttt tggtaacttg aattctacct 1992
ctcgcgaaat ctactgtaga taaaatgatt gtcatatttc cacttggaaa atgaatctcc 2052
cacggataat tgtattcatt tgaatctaag ctgtcctcca gttttaactc aactcaaacg 2112
ttttacagtt atgacagcct gttaatatga cttgtactat tttggtatta tactaataca 2172
taagagttgt acatattgtt acattcatta aatttgagaa aaattaatgt taaatacatt 2232
ttatgaacgg gccg 2246
18
535
PRT
Bovine
18
Met Arg Leu Arg Asn Gly Thr Val Ala Thr Val Leu Ala Phe Ile Thr
1 5 10 15
Ser Phe Leu Thr Leu Ser Trp Tyr Thr Thr Trp Gln Asn Gly Lys Glu
20 25 30
Lys Val Ile Ala Tyr Gln Arg Glu Phe Leu Ala Leu Lys Glu Arg Leu
35 40 45
Arg Ile Ala Glu His Arg Ile Ser Gln Arg Ser Ser Glu Leu Ser Ala
50 55 60
Ile Val Gln Gln Phe Lys Arg Val Glu Ala Glu Thr Asn Arg Ser Lys
65 70 75 80
Asp Pro Val Asn Lys Phe Ser Asp Asp Thr Leu Lys Ile Leu Lys Glu
85 90 95
Leu Thr Ser Lys Lys Ser Leu Gln Val Pro Ser Ile Tyr Tyr His Leu
100 105 110
Pro His Leu Leu Gln Asn Glu Gly Ser Leu Gln Pro Ala Val Gln Ile
115 120 125
Gly Asn Gly Arg Thr Gly Val Ser Ile Val Met Gly Ile Pro Thr Val
130 135 140
Lys Arg Glu Val Lys Ser Tyr Leu Ile Glu Thr Leu His Ser Leu Ile
145 150 155 160
Asp Asn Leu Tyr Pro Glu Glu Lys Leu Asp Cys Val Ile Val Val Phe
165 170 175
Ile Gly Glu Thr Asp Thr Asp Tyr Val Asn Gly Val Val Ala Asn Leu
180 185 190
Glu Lys Glu Phe Ser Lys Glu Ile Ser Ser Gly Leu Val Glu Ile Ile
195 200 205
Ser Pro Pro Glu Ser Tyr Tyr Pro Asp Leu Thr Asn Leu Lys Glu Thr
210 215 220
Phe Gly Asp Ser Lys Glu Arg Val Arg Trp Arg Thr Lys Gln Asn Leu
225 230 235 240
Asp Tyr Cys Phe Leu Met Met Tyr Ala Gln Glu Lys Gly Thr Tyr Tyr
245 250 255
Ile Gln Leu Glu Asp Asp Ile Ile Val Lys Gln Asn Tyr Phe Asn Thr
260 265 270
Ile Lys Asn Phe Ala Leu Gln Leu Ser Ser Glu Glu Trp Met Ile Leu
275 280 285
Glu Phe Ser Gln Leu Gly Phe Ile Gly Lys Met Phe Gln Ala Pro Asp
290 295 300
Pro Leu Leu Ile Val Glu Phe Ile Phe Met Phe Tyr Lys Glu Lys Pro
305 310 315 320
Ile Asp Trp Leu Leu Asp His Ile Leu Trp Val Lys Val Cys Asn Pro
325 330 335
Glu Lys Asp Ala Lys His Cys Asp Arg Gln Lys Ala Asn Leu Arg Ile
340 345 350
Arg Phe Arg Pro Ser Leu Phe Gln His Val Gly Leu His Ser Ser Leu
355 360 365
Thr Gly Lys Ile Gln Lys Leu Thr Asp Lys Asp Tyr Met Lys Pro Leu
370 375 380
Leu Leu Lys Ile His Val Asn Pro Pro Ala Glu Val Ser Thr Ser Leu
385 390 395 400
Lys Val Tyr Gln Gly His Thr Leu Glu Lys Thr Tyr Met Gly Glu Asp
405 410 415
Phe Phe Trp Ala Ile Thr Pro Val Ala Gly Asp Tyr Ile Leu Phe Lys
420 425 430
Phe Asp Lys Pro Val Asn Val Glu Ser Tyr Leu Phe His Ser Gly Asn
435 440 445
Gln Asp His Pro Gly Asp Ile Leu Leu Asn Thr Thr Val Glu Val Leu
450 455 460
Pro Leu Lys Ser Glu Gly Leu Asp Ile Ser Lys Glu Thr Lys Asp Lys
465 470 475 480
Arg Leu Glu Asp Gly Tyr Phe Arg Ile Gly Lys Phe Glu Asn Gly Val
485 490 495
Ala Glu Gly Met Val Asp Pro Ser Leu Asn Pro Ile Ser Ala Phe Arg
500 505 510
Leu Ser Val Ile Gln Asn Ser Ala Val Trp Ala Ile Leu Asn Glu Ile
515 520 525
His Ile Lys Lys Val Thr Asn
530 535
19
31
DNA
Artificial Sequence
Description of Artificial Sequence Primer
19
ccctcgagat gaggctccga aatggaactg t 31
20
31
DNA
Artificial Sequence
Description of Artificial Sequence Primer
20
tttctagatc agtttgtgac ttttttaata t 31
21
24
DNA
Artificial Sequence
Description of Artificial Sequence Primer
21
acgattgtgc aacagttcaa gcgt 24
22
24
DNA
Artificial Sequence
Description of Artificial Sequence Primer
22
gggagaactc caggatcatc cagt 24
23
2115
DNA
Homo sapiens
CDS
(136)..(1740)
23
gaaatgaacc tctcttattg atttttattg gcctagagcc aggagtactg cattcagttg 60
actttcaggg taaaaagaaa acagtcctgg ttgttgtcat cataaacata tggaccagtg 120
tgatggtgaa atgag atg agg ctc cgc aat gga act gta gcc act gct tta 171
Met Arg Leu Arg Asn Gly Thr Val Ala Thr Ala Leu
1 5 10
gca ttt atc act tcc ttc ctt act ttg tct tgg tat act aca tgg caa 219
Ala Phe Ile Thr Ser Phe Leu Thr Leu Ser Trp Tyr Thr Thr Trp Gln
15 20 25
aat ggg aaa gaa aaa ctg att gct tat caa cga gaa ttc ctt gct ttg 267
Asn Gly Lys Glu Lys Leu Ile Ala Tyr Gln Arg Glu Phe Leu Ala Leu
30 35 40
aaa gaa cgt ctt cga ata gct gaa cac aga atc tca cag cgc tct tct 315
Lys Glu Arg Leu Arg Ile Ala Glu His Arg Ile Ser Gln Arg Ser Ser
45 50 55 60
gaa tta aat acg att gtg caa cag ttc aag cgt gta gga gca gaa aca 363
Glu Leu Asn Thr Ile Val Gln Gln Phe Lys Arg Val Gly Ala Glu Thr
65 70 75
aat gga agt aag gat gcg ttg aat aag ttt tca gat aat acc cta aag 411
Asn Gly Ser Lys Asp Ala Leu Asn Lys Phe Ser Asp Asn Thr Leu Lys
80 85 90
ctg tta aag gag tta aca agc aaa aaa tct ctt caa gtg cca agt att 459
Leu Leu Lys Glu Leu Thr Ser Lys Lys Ser Leu Gln Val Pro Ser Ile
95 100 105
tat tat cat ttg cct cat tta ttg aaa aat gaa gga agt ctt caa cct 507
Tyr Tyr His Leu Pro His Leu Leu Lys Asn Glu Gly Ser Leu Gln Pro
110 115 120
gct gta cag att ggc aac gga aga aca gga gtt tca ata gtc atg ggc 555
Ala Val Gln Ile Gly Asn Gly Arg Thr Gly Val Ser Ile Val Met Gly
125 130 135 140
att ccc aca gtg aag aga gaa gtt aaa tct tac ctc ata gaa act ctt 603
Ile Pro Thr Val Lys Arg Glu Val Lys Ser Tyr Leu Ile Glu Thr Leu
145 150 155
cat tcc ctt att gat aac ctg tat cct gaa gag aag ttg gac tgt gtt 651
His Ser Leu Ile Asp Asn Leu Tyr Pro Glu Glu Lys Leu Asp Cys Val
160 165 170
ata gta gtc ttc ata gga gag aca gat att gat tat gta cat ggt gtt 699
Ile Val Val Phe Ile Gly Glu Thr Asp Ile Asp Tyr Val His Gly Val
175 180 185
gta gcc aac ctg gag aaa gaa ttt tct aaa gaa atc agt tct ggc ttg 747
Val Ala Asn Leu Glu Lys Glu Phe Ser Lys Glu Ile Ser Ser Gly Leu
190 195 200
gtg gaa gtc ata tca ccc cct gaa agc tat tat cct gac ttg aca aac 795
Val Glu Val Ile Ser Pro Pro Glu Ser Tyr Tyr Pro Asp Leu Thr Asn
205 210 215 220
cta aag gag aca ttt gga gac tcc aaa gaa aga gta aga tgg aga aca 843
Leu Lys Glu Thr Phe Gly Asp Ser Lys Glu Arg Val Arg Trp Arg Thr
225 230 235
aag caa aac cta gat tac tgt ttt cta atg atg tat gct caa gaa aag 891
Lys Gln Asn Leu Asp Tyr Cys Phe Leu Met Met Tyr Ala Gln Glu Lys
240 245 250
ggc ata tat tac att cag ctt gaa gat gat att att gtc aaa caa aat 939
Gly Ile Tyr Tyr Ile Gln Leu Glu Asp Asp Ile Ile Val Lys Gln Asn
255 260 265
tat ttt aat acc ata aaa aat ttt gca ctt caa ctt tct tct gag gaa 987
Tyr Phe Asn Thr Ile Lys Asn Phe Ala Leu Gln Leu Ser Ser Glu Glu
270 275 280
tgg atg att cta gag ttt tcc cag ctg ggc ttc att ggt aaa atg ttt 1035
Trp Met Ile Leu Glu Phe Ser Gln Leu Gly Phe Ile Gly Lys Met Phe
285 290 295 300
caa gcg ccg gat ctt act ctg att gta gaa ttc ata ttc atg ttt tac 1083
Gln Ala Pro Asp Leu Thr Leu Ile Val Glu Phe Ile Phe Met Phe Tyr
305 310 315
aag gag aaa ccc att gat tgg ctc ctg gac cat att ctc tgg gtg aaa 1131
Lys Glu Lys Pro Ile Asp Trp Leu Leu Asp His Ile Leu Trp Val Lys
320 325 330
gtc tgc aac cct gaa aaa gat gca aaa cat tgt gat aga cag aaa gca 1179
Val Cys Asn Pro Glu Lys Asp Ala Lys His Cys Asp Arg Gln Lys Ala
335 340 345
aat ctg cga att cgc ttc aga cct tcc ctt ttc caa cat gtt ggt ctg 1227
Asn Leu Arg Ile Arg Phe Arg Pro Ser Leu Phe Gln His Val Gly Leu
350 355 360
cac tca tca cta tca gga aaa atc caa aaa ctc acg gat aaa gat tat 1275
His Ser Ser Leu Ser Gly Lys Ile Gln Lys Leu Thr Asp Lys Asp Tyr
365 370 375 380
atg aaa cca tta ctt ctt aaa atc cat gta aac cca cct gcg gag gta 1323
Met Lys Pro Leu Leu Leu Lys Ile His Val Asn Pro Pro Ala Glu Val
385 390 395
tct act tcc ttg aag gtc tac caa ggg cat acg ctg gag aaa act tac 1371
Ser Thr Ser Leu Lys Val Tyr Gln Gly His Thr Leu Glu Lys Thr Tyr
400 405 410
atg gga gag gat ttc ttc tgg gct atc aca ccg ata gct gga gac tac 1419
Met Gly Glu Asp Phe Phe Trp Ala Ile Thr Pro Ile Ala Gly Asp Tyr
415 420 425
atc ttg ttt aaa ttt gat aaa cca gtc aat gta gaa agt tat ttg ttc 1467
Ile Leu Phe Lys Phe Asp Lys Pro Val Asn Val Glu Ser Tyr Leu Phe
430 435 440
cat agc ggc aac caa gaa cat cct gga gat att ctg cta aac aca act 1515
His Ser Gly Asn Gln Glu His Pro Gly Asp Ile Leu Leu Asn Thr Thr
445 450 455 460
gtg gaa gtt ttg cct ttt aag agt gaa ggt ttg gaa ata agc aaa gaa 1563
Val Glu Val Leu Pro Phe Lys Ser Glu Gly Leu Glu Ile Ser Lys Glu
465 470 475
acc aaa gac aaa cga tta gaa gat ggc tat ttc aga ata gga aaa ttt 1611
Thr Lys Asp Lys Arg Leu Glu Asp Gly Tyr Phe Arg Ile Gly Lys Phe
480 485 490
gag aat ggt gtt gca gaa gga atg gtg gat cca agt ctc aat ccc att 1659
Glu Asn Gly Val Ala Glu Gly Met Val Asp Pro Ser Leu Asn Pro Ile
495 500 505
tca gcc ttt cga ctt tca gtt att cag aat tct gct gtt tgg gcc att 1707
Ser Ala Phe Arg Leu Ser Val Ile Gln Asn Ser Ala Val Trp Ala Ile
510 515 520
ctt aat gag att cat att aaa aaa gcc acc aac tgatcatctg agaaaccaac 1760
Leu Asn Glu Ile His Ile Lys Lys Ala Thr Asn
525 530 535
acattttttc ctgtgaattt gttaattaaa gatagttaag catgtatctt ttttttattt 1820
ctacttgaac actacctctt gtgaagtcta ctgtagataa gacgattgtc atttccactt 1880
ggaaagtgaa tctcccataa taattgtatt tgtttgaaac taagctgtcc tcagatttta 1940
acttgactca aacatttttc aattatgaca gcctgttaat atgacttgta ctattttggt 2000
attatactaa tacataagag ttgtacatat tgttacattc tttaaatttg agaaaaacta 2060
atgttacata cattttatga agggggtact tttgaggttc acttatttta ctatt 2115
24
535
PRT
Homo sapiens
24
Met Arg Leu Arg Asn Gly Thr Val Ala Thr Ala Leu Ala Phe Ile Thr
1 5 10 15
Ser Phe Leu Thr Leu Ser Trp Tyr Thr Thr Trp Gln Asn Gly Lys Glu
20 25 30
Lys Leu Ile Ala Tyr Gln Arg Glu Phe Leu Ala Leu Lys Glu Arg Leu
35 40 45
Arg Ile Ala Glu His Arg Ile Ser Gln Arg Ser Ser Glu Leu Asn Thr
50 55 60
Ile Val Gln Gln Phe Lys Arg Val Gly Ala Glu Thr Asn Gly Ser Lys
65 70 75 80
Asp Ala Leu Asn Lys Phe Ser Asp Asn Thr Leu Lys Leu Leu Lys Glu
85 90 95
Leu Thr Ser Lys Lys Ser Leu Gln Val Pro Ser Ile Tyr Tyr His Leu
100 105 110
Pro His Leu Leu Lys Asn Glu Gly Ser Leu Gln Pro Ala Val Gln Ile
115 120 125
Gly Asn Gly Arg Thr Gly Val Ser Ile Val Met Gly Ile Pro Thr Val
130 135 140
Lys Arg Glu Val Lys Ser Tyr Leu Ile Glu Thr Leu His Ser Leu Ile
145 150 155 160
Asp Asn Leu Tyr Pro Glu Glu Lys Leu Asp Cys Val Ile Val Val Phe
165 170 175
Ile Gly Glu Thr Asp Ile Asp Tyr Val His Gly Val Val Ala Asn Leu
180 185 190
Glu Lys Glu Phe Ser Lys Glu Ile Ser Ser Gly Leu Val Glu Val Ile
195 200 205
Ser Pro Pro Glu Ser Tyr Tyr Pro Asp Leu Thr Asn Leu Lys Glu Thr
210 215 220
Phe Gly Asp Ser Lys Glu Arg Val Arg Trp Arg Thr Lys Gln Asn Leu
225 230 235 240
Asp Tyr Cys Phe Leu Met Met Tyr Ala Gln Glu Lys Gly Ile Tyr Tyr
245 250 255
Ile Gln Leu Glu Asp Asp Ile Ile Val Lys Gln Asn Tyr Phe Asn Thr
260 265 270
Ile Lys Asn Phe Ala Leu Gln Leu Ser Ser Glu Glu Trp Met Ile Leu
275 280 285
Glu Phe Ser Gln Leu Gly Phe Ile Gly Lys Met Phe Gln Ala Pro Asp
290 295 300
Leu Thr Leu Ile Val Glu Phe Ile Phe Met Phe Tyr Lys Glu Lys Pro
305 310 315 320
Ile Asp Trp Leu Leu Asp His Ile Leu Trp Val Lys Val Cys Asn Pro
325 330 335
Glu Lys Asp Ala Lys His Cys Asp Arg Gln Lys Ala Asn Leu Arg Ile
340 345 350
Arg Phe Arg Pro Ser Leu Phe Gln His Val Gly Leu His Ser Ser Leu
355 360 365
Ser Gly Lys Ile Gln Lys Leu Thr Asp Lys Asp Tyr Met Lys Pro Leu
370 375 380
Leu Leu Lys Ile His Val Asn Pro Pro Ala Glu Val Ser Thr Ser Leu
385 390 395 400
Lys Val Tyr Gln Gly His Thr Leu Glu Lys Thr Tyr Met Gly Glu Asp
405 410 415
Phe Phe Trp Ala Ile Thr Pro Ile Ala Gly Asp Tyr Ile Leu Phe Lys
420 425 430
Phe Asp Lys Pro Val Asn Val Glu Ser Tyr Leu Phe His Ser Gly Asn
435 440 445
Gln Glu His Pro Gly Asp Ile Leu Leu Asn Thr Thr Val Glu Val Leu
450 455 460
Pro Phe Lys Ser Glu Gly Leu Glu Ile Ser Lys Glu Thr Lys Asp Lys
465 470 475 480
Arg Leu Glu Asp Gly Tyr Phe Arg Ile Gly Lys Phe Glu Asn Gly Val
485 490 495
Ala Glu Gly Met Val Asp Pro Ser Leu Asn Pro Ile Ser Ala Phe Arg
500 505 510
Leu Ser Val Ile Gln Asn Ser Ala Val Trp Ala Ile Leu Asn Glu Ile
515 520 525
His Ile Lys Lys Ala Thr Asn
530 535
25
30
DNA
Artificial Sequence
Description of Artificial Sequence Primer
25
ttctcgagat gaggctccgc aatggaactg 30
26
24
DNA
Artificial Sequence
Description of Artificial Sequence Primer
26
agaaatgtgg gcttcagggc tggc 24
27
30
DNA
Artificial Sequence
Description of Artificial Sequence Primer
27
ttctcgagat gaggctccgc aatggaactg 30
28
24
DNA
Artificial Sequence
Description of Artificial Sequence Primer
28
agaaatgtgg gcttcagggc tggc 24
29
30
DNA
Artificial Sequence
Description of Artificial Sequence Primer
29
ttctcgagat gaggctccgc aatggaactg 30
30
24
DNA
Artificial Sequence
Description of Artificial Sequence Primer
30
agaaatgtgg gcttcagggc tggc 24
31
25
DNA
Artificial Sequence
Description of Artificial Sequence Primer
31
ttccatcacc tgccacacct gctgg 25
32
24
DNA
Artificial Sequence
Description of Artificial Sequence Primer
32
acaaccctca gtcagacaag gagg 24
33
24
DNA
Artificial Sequence
Description of Artificial Sequence Primer
33
acacccccag aaatgtgggc ttca 24
34
24
DNA
Artificial Sequence
Description of Artificial Sequence Primer
34
atgaccgagt cctccttctc ctgc 24
35
24
DNA
Artificial Sequence
Description of Artificial Sequence Primer
35
atgcccatca ccaccgacac tccg 24
36
1724
DNA
Homo sapiens
CDS
(43)..(1686)
36
tgcagcctcg gccccgcggg cgcccgccgc gcacccgagg ag atg agg ctc cgc 54
Met Arg Leu Arg
1
aat ggc acc ttc ctg acg ctg ctg ctc ttc tgc ctg tgc gcc ttc ctc 102
Asn Gly Thr Phe Leu Thr Leu Leu Leu Phe Cys Leu Cys Ala Phe Leu
5 10 15 20
tcg ctg tcc tgg tac gcg gca ctc agc ggc cag aaa ggc gac gtt gtg 150
Ser Leu Ser Trp Tyr Ala Ala Leu Ser Gly Gln Lys Gly Asp Val Val
25 30 35
gac gtt tac cag cgg gag ttc ctg gcg ctg cgc gat cgg ttg cac gca 198
Asp Val Tyr Gln Arg Glu Phe Leu Ala Leu Arg Asp Arg Leu His Ala
40 45 50
gct gag cag gag agc ctc aag cgc tcc aag gag ctc aac ctg gtg ctg 246
Ala Glu Gln Glu Ser Leu Lys Arg Ser Lys Glu Leu Asn Leu Val Leu
55 60 65
gac gag atc aag agg gcc gtg tca gaa agg cag gcg ctg cga gac gga 294
Asp Glu Ile Lys Arg Ala Val Ser Glu Arg Gln Ala Leu Arg Asp Gly
70 75 80
gac ggc aat cgc acc tgg ggc cgc cta aca gag gac ccc cga ttg aag 342
Asp Gly Asn Arg Thr Trp Gly Arg Leu Thr Glu Asp Pro Arg Leu Lys
85 90 95 100
ccg tgg aac ggc tca cac cgg cac gtg ctg cac ctg ccc acc gtc ttc 390
Pro Trp Asn Gly Ser His Arg His Val Leu His Leu Pro Thr Val Phe
105 110 115
cat cac ctg cca cac ctg ctg gcc aag gag agc agt ctg cag ccc gcg 438
His His Leu Pro His Leu Leu Ala Lys Glu Ser Ser Leu Gln Pro Ala
120 125 130
gtg cgc gtg ggc cag ggc cgc acc gga gtg tcg gtg gtg atg ggc atc 486
Val Arg Val Gly Gln Gly Arg Thr Gly Val Ser Val Val Met Gly Ile
135 140 145
ccg agc gtg cgg cgc gag gtg cac tcg tac ctg act gac act ctg cac 534
Pro Ser Val Arg Arg Glu Val His Ser Tyr Leu Thr Asp Thr Leu His
150 155 160
tcg ctc atc tcc gag ctg agc ccg cag gag aag gag gac tcg gtc atc 582
Ser Leu Ile Ser Glu Leu Ser Pro Gln Glu Lys Glu Asp Ser Val Ile
165 170 175 180
gtg gtg ctg atc gcc gag act gac tca cag tac act tcg gca gtg aca 630
Val Val Leu Ile Ala Glu Thr Asp Ser Gln Tyr Thr Ser Ala Val Thr
185 190 195
gag aac atc aag gcc ttg ttc ccc acg gag atc cat tct ggg ctc ctg 678
Glu Asn Ile Lys Ala Leu Phe Pro Thr Glu Ile His Ser Gly Leu Leu
200 205 210
gag gtc atc tca ccc tcc ccc cac ttc tac cct gac ttc tcc cgc ctc 726
Glu Val Ile Ser Pro Ser Pro His Phe Tyr Pro Asp Phe Ser Arg Leu
215 220 225
cga gag tcc ttt ggg gac ccc aag gag aga gtc agg tgg agg acc aaa 774
Arg Glu Ser Phe Gly Asp Pro Lys Glu Arg Val Arg Trp Arg Thr Lys
230 235 240
cag aac ctc gat tac tgc ttc ctc atg atg tac gcg cag tcc aaa ggc 822
Gln Asn Leu Asp Tyr Cys Phe Leu Met Met Tyr Ala Gln Ser Lys Gly
245 250 255 260
atc tac tac gtg cag ctg gag gat gac atc gtg gcc aag ccc aac tac 870
Ile Tyr Tyr Val Gln Leu Glu Asp Asp Ile Val Ala Lys Pro Asn Tyr
265 270 275
ctg agc acc atg aag aac ttt gca ctg cag cag cct tca gag gac tgg 918
Leu Ser Thr Met Lys Asn Phe Ala Leu Gln Gln Pro Ser Glu Asp Trp
280 285 290
atg atc ctg gag ttc tcc cag ctg ggc ttc att ggt aag atg ttc aag 966
Met Ile Leu Glu Phe Ser Gln Leu Gly Phe Ile Gly Lys Met Phe Lys
295 300 305
tcg ctg gac ctg agc ctg att gta gag ttc att ctc atg ttc tac cgg 1014
Ser Leu Asp Leu Ser Leu Ile Val Glu Phe Ile Leu Met Phe Tyr Arg
310 315 320
gac aag ccc atc gac tgg ctc ctg gac cat att ctg tgg gtg aaa gtc 1062
Asp Lys Pro Ile Asp Trp Leu Leu Asp His Ile Leu Trp Val Lys Val
325 330 335 340
tgc aac ccc gag aag gat gcg aag cac tgt gac cgg cag aaa gcc aac 1110
Cys Asn Pro Glu Lys Asp Ala Lys His Cys Asp Arg Gln Lys Ala Asn
345 350 355
ctg cgg atc cgc ttc aaa ccg tcc ctc ttc cag cac gtg ggc act cac 1158
Leu Arg Ile Arg Phe Lys Pro Ser Leu Phe Gln His Val Gly Thr His
360 365 370
tcc tcg ctg gct ggc aag atc cag aaa ctg aag gac aaa gac ttt gga 1206
Ser Ser Leu Ala Gly Lys Ile Gln Lys Leu Lys Asp Lys Asp Phe Gly
375 380 385
aag cag gcg ctg cgg aag gag cat gtg aac ccg cca gca gag gtg agc 1254
Lys Gln Ala Leu Arg Lys Glu His Val Asn Pro Pro Ala Glu Val Ser
390 395 400
acg agc ctg aag aca tac cag cac ttc acc ctg gag aaa gcc tac ctg 1302
Thr Ser Leu Lys Thr Tyr Gln His Phe Thr Leu Glu Lys Ala Tyr Leu
405 410 415 420
cgc gag gac ttc ttc tgg gcc ttc acc cct gcc gcg ggg gac ttc atc 1350
Arg Glu Asp Phe Phe Trp Ala Phe Thr Pro Ala Ala Gly Asp Phe Ile
425 430 435
cgc ttc cgc ttc ttc caa cct cta aga ctg gag cgg ttc ttc ttc cgc 1398
Arg Phe Arg Phe Phe Gln Pro Leu Arg Leu Glu Arg Phe Phe Phe Arg
440 445 450
agt ggg aac atc gag cac ccg gag gac aag ctc ttc aac acg tct gtg 1446
Ser Gly Asn Ile Glu His Pro Glu Asp Lys Leu Phe Asn Thr Ser Val
455 460 465
gag gtg ctg ccc ttc gac aac cct cag tca gac aag gag gcc ctg cag 1494
Glu Val Leu Pro Phe Asp Asn Pro Gln Ser Asp Lys Glu Ala Leu Gln
470 475 480
gag ggc cgc acc gcc acc ctc cgg tac cct cgg agc ccc gac ggc tac 1542
Glu Gly Arg Thr Ala Thr Leu Arg Tyr Pro Arg Ser Pro Asp Gly Tyr
485 490 495 500
ctc cag atc ggc tcc ttc tac aag gga gtg gca gag gga gag gtg gac 1590
Leu Gln Ile Gly Ser Phe Tyr Lys Gly Val Ala Glu Gly Glu Val Asp
505 510 515
cca gcc ttc ggc cct ctg gaa gca ctg cgc ctc tcg atc cag acg gac 1638
Pro Ala Phe Gly Pro Leu Glu Ala Leu Arg Leu Ser Ile Gln Thr Asp
520 525 530
tcc cct gtg tgg gtg att ctg agc gag atc ttc ctg aaa aag gcc gac 1686
Ser Pro Val Trp Val Ile Leu Ser Glu Ile Phe Leu Lys Lys Ala Asp
535 540 545
taagctgcgg gcttctgagg gtaccctgtg gccagccc 1724
37
548
PRT
Homo sapiens
37
Met Arg Leu Arg Asn Gly Thr Phe Leu Thr Leu Leu Leu Phe Cys Leu
1 5 10 15
Cys Ala Phe Leu Ser Leu Ser Trp Tyr Ala Ala Leu Ser Gly Gln Lys
20 25 30
Gly Asp Val Val Asp Val Tyr Gln Arg Glu Phe Leu Ala Leu Arg Asp
35 40 45
Arg Leu His Ala Ala Glu Gln Glu Ser Leu Lys Arg Ser Lys Glu Leu
50 55 60
Asn Leu Val Leu Asp Glu Ile Lys Arg Ala Val Ser Glu Arg Gln Ala
65 70 75 80
Leu Arg Asp Gly Asp Gly Asn Arg Thr Trp Gly Arg Leu Thr Glu Asp
85 90 95
Pro Arg Leu Lys Pro Trp Asn Gly Ser His Arg His Val Leu His Leu
100 105 110
Pro Thr Val Phe His His Leu Pro His Leu Leu Ala Lys Glu Ser Ser
115 120 125
Leu Gln Pro Ala Val Arg Val Gly Gln Gly Arg Thr Gly Val Ser Val
130 135 140
Val Met Gly Ile Pro Ser Val Arg Arg Glu Val His Ser Tyr Leu Thr
145 150 155 160
Asp Thr Leu His Ser Leu Ile Ser Glu Leu Ser Pro Gln Glu Lys Glu
165 170 175
Asp Ser Val Ile Val Val Leu Ile Ala Glu Thr Asp Ser Gln Tyr Thr
180 185 190
Ser Ala Val Thr Glu Asn Ile Lys Ala Leu Phe Pro Thr Glu Ile His
195 200 205
Ser Gly Leu Leu Glu Val Ile Ser Pro Ser Pro His Phe Tyr Pro Asp
210 215 220
Phe Ser Arg Leu Arg Glu Ser Phe Gly Asp Pro Lys Glu Arg Val Arg
225 230 235 240
Trp Arg Thr Lys Gln Asn Leu Asp Tyr Cys Phe Leu Met Met Tyr Ala
245 250 255
Gln Ser Lys Gly Ile Tyr Tyr Val Gln Leu Glu Asp Asp Ile Val Ala
260 265 270
Lys Pro Asn Tyr Leu Ser Thr Met Lys Asn Phe Ala Leu Gln Gln Pro
275 280 285
Ser Glu Asp Trp Met Ile Leu Glu Phe Ser Gln Leu Gly Phe Ile Gly
290 295 300
Lys Met Phe Lys Ser Leu Asp Leu Ser Leu Ile Val Glu Phe Ile Leu
305 310 315 320
Met Phe Tyr Arg Asp Lys Pro Ile Asp Trp Leu Leu Asp His Ile Leu
325 330 335
Trp Val Lys Val Cys Asn Pro Glu Lys Asp Ala Lys His Cys Asp Arg
340 345 350
Gln Lys Ala Asn Leu Arg Ile Arg Phe Lys Pro Ser Leu Phe Gln His
355 360 365
Val Gly Thr His Ser Ser Leu Ala Gly Lys Ile Gln Lys Leu Lys Asp
370 375 380
Lys Asp Phe Gly Lys Gln Ala Leu Arg Lys Glu His Val Asn Pro Pro
385 390 395 400
Ala Glu Val Ser Thr Ser Leu Lys Thr Tyr Gln His Phe Thr Leu Glu
405 410 415
Lys Ala Tyr Leu Arg Glu Asp Phe Phe Trp Ala Phe Thr Pro Ala Ala
420 425 430
Gly Asp Phe Ile Arg Phe Arg Phe Phe Gln Pro Leu Arg Leu Glu Arg
435 440 445
Phe Phe Phe Arg Ser Gly Asn Ile Glu His Pro Glu Asp Lys Leu Phe
450 455 460
Asn Thr Ser Val Glu Val Leu Pro Phe Asp Asn Pro Gln Ser Asp Lys
465 470 475 480
Glu Ala Leu Gln Glu Gly Arg Thr Ala Thr Leu Arg Tyr Pro Arg Ser
485 490 495
Pro Asp Gly Tyr Leu Gln Ile Gly Ser Phe Tyr Lys Gly Val Ala Glu
500 505 510
Gly Glu Val Asp Pro Ala Phe Gly Pro Leu Glu Ala Leu Arg Leu Ser
515 520 525
Ile Gln Thr Asp Ser Pro Val Trp Val Ile Leu Ser Glu Ile Phe Leu
530 535 540
Lys Lys Ala Asp
545
38
29
DNA
Artificial Sequence
Description of Artificial Sequence Primer
38
ttctcgagga gatgaggctc cgcaatggc 29
39
29
DNA
Artificial Sequence
Description of Artificial Sequence Primer
39
aatctagaaa tgtgggcttc agggctggc 29
40
28
DNA
Artificial Sequence
Description of Artificial Sequence Primer
40
ccctcgagat gggggtgcac gaatgtcc 28
41
48
DNA
Artificial Sequence
Description of Artificial Sequence Primer
41
ctttttgctt gttaactcct ttagtattgg ggcgcccagg actgggag 48
42
48
DNA
Artificial Sequence
Description of Artificial Sequence Primer
42
ctcccagtcc tgggcgcccc aatactaaag gagttaacaa gcaaaaag 48
43
44
DNA
Artificial Sequence
Description of Artificial Sequence Primer
43
cttccttcat tttgcaataa atgaggggcg cccaggactg ggag 44
44
43
DNA
Artificial Sequence
Description of Artificial Sequence Primer
44
atttaacttc tctcttcact gtaggggcgc ccaggactgg gag 43
45
44
DNA
Artificial Sequence
Description of Artificial Sequence Primer
45
ctcccagtcc tgggcgcccc tcatttattg caaaatgaag gaag 44
46
43
DNA
Artificial Sequence
Description of Artificial Sequence Primer
46
ctcccagtcc tgggcgcccc tacagtgaag agagaagtta aat 43
47
26
DNA
Artificial Sequence
Description of Artificial Sequence Primer
47
ttctagaatc acccttccgc aacacc 26
48
28
DNA
Artificial Sequence
Description of Artificial Sequence Primer
48
ttctagaatc aaggcagaac ttccaccg 28
49
38
DNA
Artificial Sequence
Description of Artificial Sequence Primer
49
ttctagaatc atttaaatag gatgtagtct ccagctac 38
50
34
DNA
Artificial Sequence
Description of Artificial Sequence Primer
50
ttctagaatc atggtttcat gtaatcttta tccg 34
51
19
DNA
Artificial Sequence
Description of Artificial Sequence Primer
51
ttcatgaggc tccgaaatg 19
52
29
DNA
Artificial Sequence
Description of Artificial Sequence Primer
52
caagcttcag tttgtgactt ttttaatat 29
53
27
DNA
Artificial Sequence
Description of Artificial Sequence Primer
53
tcatgatact aaaggagtta acaagca 27
54
51
DNA
Artificial Sequence
Description of Artificial Sequence Primer
54
caagcttcag tggtggtggt ggtggtggtt tgtgactttt ttaatatgga t 51
55
18
DNA
Artificial Sequence
Description of Artificial Sequence Primer
55
caagcttcag tggtggtg 18
56
32
DNA
Artificial Sequence
Description of Artificial Sequence Primer
56
tcatgatatt aaaggagtta acaagcaaaa aa 32
57
51
DNA
Artificial Sequence
Description of Artificial Sequence Primer
57
caagcttcag tggtggtggt ggtggtggtt ggtggctttt ttaatatgaa t 51
58
49
DNA
Artificial Sequence
Description of Artificial Sequence Primer
58
caagcttcag tggtggtggt ggtggtgtgg tttcatataa tctttatcc 49
59
19
DNA
Artificial Sequence
Description of Artificial Sequence Primer
59
ttcatgaggc tccgcaatg 19
60
18
DNA
Artificial Sequence
Description of Artificial Sequence Primer
60
caggttgagc tccttgga 18
61
18
DNA
Artificial Sequence
Description of Artificial Sequence Primer
61
tccaaggagc tcaacctg 18
62
44
DNA
Artificial Sequence
Description of Artificial Sequence Primer
62
caagcttcag tggtggtggt ggtggtggtc ggcctttttc agga 44
63
23
DNA
Artificial Sequence
Description of Artificial Sequence Primer
63
cccatgggcc gcctaacaga gga 23